






Documental sobre el proyecto MK-Ultra, un proyecto que ha implicado a muchos científicos desde los años 50 del siglo XX.
söndag 20 juni 2010
torsdag 6 augusti 2009
EFECTO DE AUDICIÓN DE MICROONDAS
El efecto de audición de microondas (microwave auditory effect)también conocido por el efecto Frey (Frey effect) es un efecto detectado por primera vez en la segunda guerra mundial por personas que trabajaban en las cercanías de los transpondedores (repetidores de radio especiales) de radar y consiste en sonidos audibles inducidos por impulsos de frecuencias de microondas. Los sonidos se generan directamente en el interior de la cabeza humana sin la necesidad de cualquier dispositivo electrónico.
Más tarde el neurocientífico americano Allan H. Frey estudió este fenómeno y fue el primero en publicar (Journal of Applied Physiology, vol. 17, páginas 689-692, 1962) información sobre la naturaleza del efecto auditivo de microondas, este efecto es lo por tanto, también conocido como el efecto Frey.
El Dr. Don R. Justesen publicó "Microondas y comportamiento" (Microwaves and Behavior) en el American Psychologist (Volumen 30, de marzo de 1975, Número 3).
Las investigaciones realizadas por la NASA en la década de 1970 mostró que este efecto se produce como consecuencia de la expansión térmica de las partes del oído humano en torno a la cóclea, incluso a baja densidad de potencia. Más tarde, se encontró la señal de modulación para producir sonidos o palabras que al parecer se originan intracranealmente. Se estudió para su posible uso en las comunicaciones, pero no se ha desarrollado debido a los posibles efectos biológicos de la peligrosa radiación de microondas. Investigación similar realizada en la URSS estudió su uso en armas no letales.

La existencia de armas no letales que explotan el efecto auditivo de microondas se decretó como "Secreto clasificado" en los EE.UU. a partir de (al menos) 1998, la desclasificación fué el 6 de diciembre de 2006 de "Bioeffects of Selected Non-Lethal Weaponry" en respuesta a una solicitud de FOIA.
FOIA (Freedom of Information Act)

ARTÍCULOS RELACIONADOS:
VOCES EN LA CABEZA
Artículo de la BBC sobre el asesinato de la Ministra sueca A. Lind.
Artículo de la BBC
MIND GAMES BY WASHINGTON POST
Más tarde el neurocientífico americano Allan H. Frey estudió este fenómeno y fue el primero en publicar (Journal of Applied Physiology, vol. 17, páginas 689-692, 1962) información sobre la naturaleza del efecto auditivo de microondas, este efecto es lo por tanto, también conocido como el efecto Frey.
El Dr. Don R. Justesen publicó "Microondas y comportamiento" (Microwaves and Behavior) en el American Psychologist (Volumen 30, de marzo de 1975, Número 3).
Las investigaciones realizadas por la NASA en la década de 1970 mostró que este efecto se produce como consecuencia de la expansión térmica de las partes del oído humano en torno a la cóclea, incluso a baja densidad de potencia. Más tarde, se encontró la señal de modulación para producir sonidos o palabras que al parecer se originan intracranealmente. Se estudió para su posible uso en las comunicaciones, pero no se ha desarrollado debido a los posibles efectos biológicos de la peligrosa radiación de microondas. Investigación similar realizada en la URSS estudió su uso en armas no letales.

La existencia de armas no letales que explotan el efecto auditivo de microondas se decretó como "Secreto clasificado" en los EE.UU. a partir de (al menos) 1998, la desclasificación fué el 6 de diciembre de 2006 de "Bioeffects of Selected Non-Lethal Weaponry" en respuesta a una solicitud de FOIA.
FOIA (Freedom of Information Act)

ARTÍCULOS RELACIONADOS:
VOCES EN LA CABEZA
Artículo de la BBC sobre el asesinato de la Ministra sueca A. Lind.
Artículo de la BBC
MIND GAMES BY WASHINGTON POST
MICROONDAS Y CONDUCTA
EL SIGUIENTE ENLACE CONTIENE IMPORTANTE DOCUMENTACIÓN Y REFERENCIAS DE PUBLICACIONES CIENTÍFICAS SOBRE EL TEMA MICROONDAS Y CONDUCTA. LAS REFERENCIAS SON ANTIGUAS, DEMOSTRANDOSE QUE LA TECNOLOGÍA NECESARIA PARA PRODUCIR "VOCES EN LA CABEZA" EXISTE DESDE HACE AL MENOS 40 AÑOS PROBABLEMENTE MÁS.
MICROONDAS Y CONDUCTA
ESPERO QUE LOS QUE ACCEDAN A ESTE ENLACE, PUEDAN COMPRENDER LA IMPORTANTE INFORMACIÓN QUE SE ESTÁ OCULTANDO A LA SOCIEDAD Y EL PELIGRO QUE SUPONE EL USO CLANDESTINO DE ESTAS TECNOLOGÍAS CON SERES HUMANOS.
MICROONDAS Y CONDUCTA
ESPERO QUE LOS QUE ACCEDAN A ESTE ENLACE, PUEDAN COMPRENDER LA IMPORTANTE INFORMACIÓN QUE SE ESTÁ OCULTANDO A LA SOCIEDAD Y EL PELIGRO QUE SUPONE EL USO CLANDESTINO DE ESTAS TECNOLOGÍAS CON SERES HUMANOS.
Referencia blibliográfica
Los siguientes documentos son una copia de la revista American Psychologist. The Official Journal of the American Psychological Association de Marzo de 1975. En este artículo se explica el fundamento científico y tecnológico de la energía dirigida responsable de "oir voces en la cabeza". Esto demuestra que ya a mediados de los años 70, la tecnología existía y era conocida en los medios científicos. Las partes mas relevantes del artículo están marcadas. Por favor, pinchar las imágenes para agrandarlas.








måndag 27 juli 2009
Control mental del ordenador con BrainGate
BrainGate es una nueva técnica destinada a ayudar a las personas con daños en la médula espinal. Consiste en la implantación de un chip de pocos milímetros en el cráneo del paciente que capta los impulsos eléctricos de su cerebro. Un ordenador se encarga de interpretar dichos impulsos como acciones.
Este informativo demuestra que la tecnología para descodificar los las perturbaciones electromagnéticas producidas por el cerebro al pensar, es una realidad. Pero para conocer la historía de estos avances en neurociencias leer el artículo de Washington Post : WASHINGTON POST: MIND GAMES
Realmente la tecnología para descodificar las perturbaciones electromagnéticas producidas por el cerebro existe desde hace décadas. La novedad es que ahora se está aplicando en medicina con la espectativa de incorporar estas tecnologías en la sanidad pública.
Este informativo demuestra que la tecnología para descodificar los las perturbaciones electromagnéticas producidas por el cerebro al pensar, es una realidad. Pero para conocer la historía de estos avances en neurociencias leer el artículo de Washington Post : WASHINGTON POST: MIND GAMES
Realmente la tecnología para descodificar las perturbaciones electromagnéticas producidas por el cerebro existe desde hace décadas. La novedad es que ahora se está aplicando en medicina con la espectativa de incorporar estas tecnologías en la sanidad pública.
onsdag 22 juli 2009
COMPRENSIÓN DEL SISTEMA NERVIOSO
A continuación un artículo titulado "Comprensión del sistema nervioso: Una perspectiva desde la ingeniería" de Sid Deutsch and Alice Deutsch
Source:Understanding the nervous system An engineering perspective, 1993 by Sid Deutsch and Alice Deutsch
In the extracts below the author mentions that some people fear that, somehow, their minds may be read from EEG signal recordings and then explains that this cannot be the case since the EEG signals only reflect some kind of crude average of the many unresolved cortical potentials.
I believe this is a representative view among the wellinformed and this is why I quoted it. Do note however, that the author in no way denies that by measuring those cortical potentials directly it becomes an entirely different mater.
There is also a passage stating that "..a dream remains the private property of its originator", which is credible in the context of EEG measurements, while this privacy does not extend to the context of direct connections to the nervous system.
The preface was quoted to give the reader an idea of what to expect, when actually consulting this source.
The title of the book is revealing and it is the kind of information an engineer, who has no access to classified information, will be looking for when contemplating the basic electronic aspects connected with cybernetic applications.
There is also a joking remark in the text advicing not to let a Ph D attach electrodes to a patient or anybody else.
Although it is stated jokingly I fear that in the context of these home pages, it might be interpreted to mean something else. Since I am in favor of legitimate uses of all kinds of advanced technology for the benefit of mankind, I feel compelled to state my view that as long as there is a proper communication with the patient, there is no reason to block the use of any particular type of useful technology.
There is no reason to add to the irrational fears among the public.
What people should be warned about is the genuine danger of secret uses of technology for obscure purposes.
--------------------------------------------------------------------------------
Preface
In 1987 a book that I wrote with Evangelia M-Tzanakou of Rutgers University, Neuroelectric Systems, was published by New York University Press. Sometime before this manuscript was completed it occurred to me that one ought to write a book, about the nervous system, for the lay person.
Of course, many books of this type have been written by life scientists, but a book written by a biomedical engineer would be different. Although some of my relatives and best friends are life scientists, there are two cultures. The world in general and the nervous system in particular look quite different to an engineer than they do to a life scientist.
I proceeded to generate a manuscript without equations (or, at least, nothing more complicated than Rate x Time = Distance). It was a challenge, a very enjoyable challenge, to explain the nervous system from an engineering perspective to the intelligent lay person. Alas, the manuscript was unpublishable. Even a literary agent did not help. Lay persons were undoubtedly interested in the nervous system, but not as seen through the eyes of an engineer.
Undaunted, I enlisted the aid of my daughter Alice, who is one of the life scientists that has a Ph.D. in biology. Alice made a semi-infinite number of changes, adding and subtracting so much material that it was only fair to call her a co-author. Unfortunately, this fooled nobody. The revised manuscript remained too heavily flavored by an engineering viewpoint.
Finally, I sent the manuscript to a publisher of engineering books - namely, IEEE PRESS. The PRESS's reviewers said, "Put the equations back, and you will have something that is viable!" I went even further-I cheerfully gathered up my favorite home work and exam questions, the residue of 25 years of teaching neuroelectric systems, and added Problems at the end of each chapter. (This includes answers to those exam gems that were formerly carefully guarded.)
One final change was made. One of the reviewers, Murray Eden of NIH, opined that most of the mathematics should be presented in appendix form at the end of each pertinent chapter. I am grateful to Dr. Eden for this excellent suggestion.
As the chapter headings indicate, the book begins with the basic components of the nervous system-with sensory receptors, neurons and their dendrites, and skeletal muscle circuits. This is followed by the more complicated auditory and visual systems. Finally, the climax - the brain - is considered. The 12 chapters correspond to a three-credit biomedical engineering course on the nervous system.
What is the engineering perspective? It is one in which each physiological system is simplified until it can be analyzed mathematically, followed by presentation as a simplified model.
The material in the book has been shaped by a myriad of students and colleagues-at the Polytechnic Institute of Brooklyn (now Polytechnic University) until 1972; at Rutgers University from 1972 to 1979; at Tel Aviv University from 1979 to 1983; and at the University of South Florida (as a "Visitor") since 1983. The manuscript was also modified in accordance with the helpful comments of some six "anonymous" reviewers, in addition to those of Dr. Eden.
I am thankful to the people of USF for their cooperation-especially Tom Smith and Dr. Elias Stefanakos of the Electrical Engineering Department, and Dr. Michael Kovac, Dean of Engineering. Special thanks are also due to the editorial staff of IEEE PRESS, notably Executive Editor Dudley Kay, Production Supervisor Denise Gannon, and Production Editor Karen Miller, without whose expertise this project could not have come to fruition.
Above all, Alice and I are grateful to Ruth, mother and wife, who was one of the "intelligent lay persons" who undertook to read the first few chapters of the first manuscript.
Sid Deutsch Sarasota, FL
Ever since I can remember, my father always answered my "why" and "how" questions about the world. No matter what he was doing, he always had time to give me a scientific explanation for some phenomenon of our environment. No question was ever answered by that oft-quoted phrase: "Look it up!" I still remember his description of Einstein's theory of relativity that we discussed when I was a preteen. When he came to me with the suggestion that I broaden his engineering interpretation of the nervous system, I was delighted. I saw this as my chance, in turn, to help him understand the whys and hows of the nervous system from the biologist's point of view. Biology relies so completely on experimentally proven facts. In contrast, I saw the book as a series of gross approximations about various aspects of the nervous system which had as starting points these facts. It was fascinating to me that in this way - using logical mathematical expressions and formulas - one could attempt to explain biological data. I suppose it is the dialectic of starting with some basic observations, developing a theory - in this case often a mathematical model - to explain the facts, and from this, designing better experiments so as to learn yet more facts which in turn will lead to a better mathematical description or model, and so on, until the "absolute" truth about the nervous system is known. It is my hope that this book will serve as a unique point of view about the nervous system that can stimulate both new research and new theories. I also hope that it can lead to a greater understanding of the nervous system by being valuable to both the bioengineer and the neurophysiologist.
Alice Deutsch New York, NY
- - -
1 - 3 (p 13)
...A "thought" that lasts for several seconds is the outcome of millions of cortical potentials chasing around inside the brain. A pair of electroencephalographic (EEG) electrodes attached to the head shows an unending stream of irregular discharges. It does not do much good to "think simple" during an EEG examination; the electrical signal remains incomprehensible. (Its magnitude is around 10 mV 1.)
[S. D. writes: "I once was approached by a patient who insisted that certain people were 'listening in' to his inner thoughts by analyzing his EEG signals, and that they were able to do this without even using electrodes. Alas, after many years of sophisticated monitoring of brain waves, one can only conclude that 'millions of cortical potentials are chasing around inside the brain'; the nature of the signal radically changes during trauma or epileptic seizures and during sleep, but a dream remains the private property of its originator."]
One final analogy may be helpful. Imagine that you are in a satellite looking down upon a large city. You cannot see individual people, of course. But if thousands of bodies march on the Capitol building, you may be able to see a blob, the integrated outcome of many individual images. Similarly, a pair of scalp electrodes can not "see" individual brain potentials on the other side of relatively thick skull bone. What it does see is the momentary movement of thousands of signals that happen to be "marching" in the same direction, but this direction changes many times in one second.
- - -
12-3 Electroencephalographic (EEG) Signals
In 1924, Hans Berger connected two electrodes (small, round, metal disks) to a patient's scalp and detected a feeble current by using a very delicate galvanometer. Berger thereby opened up a window on the brain. As electronic amplifiers and recording equipment came into common use, the brain signal - called an electroencephalogram (EEG) - became a major clinical tool of the neurologist [J. D. Bronzino, 1984; R. Elul, 1972; A. S. Gevins, 1984; A. S. Gevins et al., 1975; K. A. Kooi, R. P. Tucker & R. E. Marshall, 1978; M. Matonsek et al., 1967; G. Pfurtscheller and R. Cooper, 1975; A. Remond, 1971; E. Vaadia, H. Bergman & M. Abeles, 1989].
Five EEG records taken from a normal individual are shown in Fig. 12-4 [H. H. Jasper, 1941]. The time scale is given by the 1-s line at the bottom. The voltage scales are given by the 50- mV 1 lines at the right side. For this type of recording, the "hot" electrode is usually located on the back of the head, opposite the striate cortex, and the other on a neutral point such as an ear lobe. In order to get a good recording, one must of course make good contact with the skin. It is not necessary to shave the patient's head, but skin oil should be removed with a solvent. A conducting paste is applied to the electrode, which is held by a suitable strap or cap against the scalp. The neutral ear-lobe electrode can be held in place with a simple clip.
[S.D. writes: "When I was at the Rutgers Medical School in New Jersey, I designed equipment for telemetering 15 simultaneous EEGs to a receiver in an adjoining room, via FM, so that the animal or human patient could be

Fig. 12-4 Electroencephalographic recordings taken from a normal individual. For this type of recording, the "hot" electrode is usually located on the back of the head, the other on a neutral point such as an ear lobe. In the "relaxed" waveform the patient's eyes are closed. Notice the time and voltage scales: the 1-s line at the bottom and 50-mV 1 lines at the right side. (From Jasper, Epilepsy and cerebral localization, Thomas, 1941.)
free to move about. In the first test of the equipment I volunteered to be the patient. My neurologist friend and colleague insisted that the normal solvent treatment was inadequate for my oily skin, and proceeded to sandpaper my ear lobe until it bled. The recording was normal, but I was the buff of jokes for the next week or two about how my ear became bitten. The moral of the story-don't let a Ph.D. connect EEG electrodes to a patient or anybody else."]
The ear-lobe electrode does not pick up appreciable signals because it is relatively far away from electrical sources such as brain cells and muscle fibers; in fact, that is why it is considered to be a neutral point. The other electrode only "sees" the electrical activity in its immediate vicinity. To begin with, because of the thickness of scalp, muscles, skull bone, and mem- brane surrounding the brain, the electrode is around 1 cm away from the cerebral cortex. Signals from neurons deeper in the brain are rapidly attenuated as distance increases. The traces of Fig. 12-4 therefore reveal local happenings. The voltage from an individual neuron is far too small to show up on an electrode l cm away. What we are seeing are the voltages from thousands of neurons whose action or graded potentials happen to be headed in the same direction at the same time. It is somewhat like being up in a satellite and trying to see individuals on earth. If everybody in Times Square suddenly headed north, say, you could possibly see the flow in one direction. Normally, however, people moving one way cancel out the people moving in the opposite direction.
In the first waveform of Fig. 12-4, the patient is "excited" (but not agitated) in the sense that he (or she) is looking at the people in the room and listening to their conversations. To avoid interference from the APs that elicit muscle contraction, the patient is not talking or moving about. We see that the EEG is a weak (10 mV 1 ), chaotic signal. From the frequencies present in the trace, we can conclude that it is not derived from high-speed (100 meters/second (m/s» APs, which would induce relatively high frequencies as they fly by. It is not even derived from slow 1 m/s APs. The frequencies are reminiscent of the 0.1-m/s progression of a graded potential of maximum amplitude 20 mV as it is conducted along a dendrite: its neuron integrates many inputs to generate its own graded potential, and the latter is "leisurely"
transmitted to another dendrite via an electrical gap junction or via a chemical agent that diffuses across a synaptic junction.
The fear voiced by some individuals that the neurologist can read their "inner thoughts" is without foundation. Certain regularities can be observed if the person is looking at, say, a pattern of vertical stripes, but the same thought repeated over and over again merely results in a random, nonrepetitive and meaningless EEG pattern [Skeptical Jnquirer].
In the second waveform, which is "relaxed," the patient' s eyes are closed. This state gives rise to a relatively strong, nonsteady oscillation at around 10 Hz, known as the alpha rhythm [H. T. Castello, 1983; B. H. Jansen, 1985; J. G. Okyere, P. Y. Ktonas & J. S. Meyer, 1986]. The origin of the alpha rhythm is controversial, but it is probably connected to vision because it is strongest in the vicinity of the striate cortex. Especially mysterious is the fact that it vanishes when the patient opens his or her eyes and looks at the world about. A hypothetical explanation is offered at the end of this section.
As the patient progresses (or retrogresses) from relaxed to drowsy to asleep to deep sleep, radical changes in frequency and amplitude take place. "Deep sleep" displays slow waves, only a few cycles per second, of large amplitude (100 mV 1). The large amplitude indicates that entire sections of tissue beneath the electrode are undergoing the same electrochemical changes in unison. The slow waves hint at the relatively slow transport of jons. Perhaps the EEG is telling us that the debris following many hours of wakefulness is being removed and "normal" material is being replenished. Altematively, in Chapter II, it is suggested that self-organization lakes place during "quiet" periods such as sleep. These are relatively slow processes; the human cycle consists of 16 hours of alertness followed by eight hours of sleep (not all of it, of course, as "deep sleep") [A. R. Morrison, 1983].
The clinical value of the EEG comes from the fact that an abnormal waveform is correlated with abnormalcy underneath the electrode [N. I. Bachen, 1986; D. G. Childers et al., 1982; C. D. McGillem, J. I. Aunon & K.-B. Vu, 1985; N. H. Morgan and A. S. Gevins, 1986; G. Pfurtscheller,
Source:Understanding the nervous system An engineering perspective, 1993 by Sid Deutsch and Alice Deutsch
In the extracts below the author mentions that some people fear that, somehow, their minds may be read from EEG signal recordings and then explains that this cannot be the case since the EEG signals only reflect some kind of crude average of the many unresolved cortical potentials.
I believe this is a representative view among the wellinformed and this is why I quoted it. Do note however, that the author in no way denies that by measuring those cortical potentials directly it becomes an entirely different mater.
There is also a passage stating that "..a dream remains the private property of its originator", which is credible in the context of EEG measurements, while this privacy does not extend to the context of direct connections to the nervous system.
The preface was quoted to give the reader an idea of what to expect, when actually consulting this source.
The title of the book is revealing and it is the kind of information an engineer, who has no access to classified information, will be looking for when contemplating the basic electronic aspects connected with cybernetic applications.
There is also a joking remark in the text advicing not to let a Ph D attach electrodes to a patient or anybody else.
Although it is stated jokingly I fear that in the context of these home pages, it might be interpreted to mean something else. Since I am in favor of legitimate uses of all kinds of advanced technology for the benefit of mankind, I feel compelled to state my view that as long as there is a proper communication with the patient, there is no reason to block the use of any particular type of useful technology.
There is no reason to add to the irrational fears among the public.
What people should be warned about is the genuine danger of secret uses of technology for obscure purposes.
--------------------------------------------------------------------------------
Preface
In 1987 a book that I wrote with Evangelia M-Tzanakou of Rutgers University, Neuroelectric Systems, was published by New York University Press. Sometime before this manuscript was completed it occurred to me that one ought to write a book, about the nervous system, for the lay person.
Of course, many books of this type have been written by life scientists, but a book written by a biomedical engineer would be different. Although some of my relatives and best friends are life scientists, there are two cultures. The world in general and the nervous system in particular look quite different to an engineer than they do to a life scientist.
I proceeded to generate a manuscript without equations (or, at least, nothing more complicated than Rate x Time = Distance). It was a challenge, a very enjoyable challenge, to explain the nervous system from an engineering perspective to the intelligent lay person. Alas, the manuscript was unpublishable. Even a literary agent did not help. Lay persons were undoubtedly interested in the nervous system, but not as seen through the eyes of an engineer.
Undaunted, I enlisted the aid of my daughter Alice, who is one of the life scientists that has a Ph.D. in biology. Alice made a semi-infinite number of changes, adding and subtracting so much material that it was only fair to call her a co-author. Unfortunately, this fooled nobody. The revised manuscript remained too heavily flavored by an engineering viewpoint.
Finally, I sent the manuscript to a publisher of engineering books - namely, IEEE PRESS. The PRESS's reviewers said, "Put the equations back, and you will have something that is viable!" I went even further-I cheerfully gathered up my favorite home work and exam questions, the residue of 25 years of teaching neuroelectric systems, and added Problems at the end of each chapter. (This includes answers to those exam gems that were formerly carefully guarded.)
One final change was made. One of the reviewers, Murray Eden of NIH, opined that most of the mathematics should be presented in appendix form at the end of each pertinent chapter. I am grateful to Dr. Eden for this excellent suggestion.
As the chapter headings indicate, the book begins with the basic components of the nervous system-with sensory receptors, neurons and their dendrites, and skeletal muscle circuits. This is followed by the more complicated auditory and visual systems. Finally, the climax - the brain - is considered. The 12 chapters correspond to a three-credit biomedical engineering course on the nervous system.
What is the engineering perspective? It is one in which each physiological system is simplified until it can be analyzed mathematically, followed by presentation as a simplified model.
The material in the book has been shaped by a myriad of students and colleagues-at the Polytechnic Institute of Brooklyn (now Polytechnic University) until 1972; at Rutgers University from 1972 to 1979; at Tel Aviv University from 1979 to 1983; and at the University of South Florida (as a "Visitor") since 1983. The manuscript was also modified in accordance with the helpful comments of some six "anonymous" reviewers, in addition to those of Dr. Eden.
I am thankful to the people of USF for their cooperation-especially Tom Smith and Dr. Elias Stefanakos of the Electrical Engineering Department, and Dr. Michael Kovac, Dean of Engineering. Special thanks are also due to the editorial staff of IEEE PRESS, notably Executive Editor Dudley Kay, Production Supervisor Denise Gannon, and Production Editor Karen Miller, without whose expertise this project could not have come to fruition.
Above all, Alice and I are grateful to Ruth, mother and wife, who was one of the "intelligent lay persons" who undertook to read the first few chapters of the first manuscript.
Sid Deutsch Sarasota, FL
Ever since I can remember, my father always answered my "why" and "how" questions about the world. No matter what he was doing, he always had time to give me a scientific explanation for some phenomenon of our environment. No question was ever answered by that oft-quoted phrase: "Look it up!" I still remember his description of Einstein's theory of relativity that we discussed when I was a preteen. When he came to me with the suggestion that I broaden his engineering interpretation of the nervous system, I was delighted. I saw this as my chance, in turn, to help him understand the whys and hows of the nervous system from the biologist's point of view. Biology relies so completely on experimentally proven facts. In contrast, I saw the book as a series of gross approximations about various aspects of the nervous system which had as starting points these facts. It was fascinating to me that in this way - using logical mathematical expressions and formulas - one could attempt to explain biological data. I suppose it is the dialectic of starting with some basic observations, developing a theory - in this case often a mathematical model - to explain the facts, and from this, designing better experiments so as to learn yet more facts which in turn will lead to a better mathematical description or model, and so on, until the "absolute" truth about the nervous system is known. It is my hope that this book will serve as a unique point of view about the nervous system that can stimulate both new research and new theories. I also hope that it can lead to a greater understanding of the nervous system by being valuable to both the bioengineer and the neurophysiologist.
Alice Deutsch New York, NY
- - -
1 - 3 (p 13)
...A "thought" that lasts for several seconds is the outcome of millions of cortical potentials chasing around inside the brain. A pair of electroencephalographic (EEG) electrodes attached to the head shows an unending stream of irregular discharges. It does not do much good to "think simple" during an EEG examination; the electrical signal remains incomprehensible. (Its magnitude is around 10 mV 1.)
[S. D. writes: "I once was approached by a patient who insisted that certain people were 'listening in' to his inner thoughts by analyzing his EEG signals, and that they were able to do this without even using electrodes. Alas, after many years of sophisticated monitoring of brain waves, one can only conclude that 'millions of cortical potentials are chasing around inside the brain'; the nature of the signal radically changes during trauma or epileptic seizures and during sleep, but a dream remains the private property of its originator."]
One final analogy may be helpful. Imagine that you are in a satellite looking down upon a large city. You cannot see individual people, of course. But if thousands of bodies march on the Capitol building, you may be able to see a blob, the integrated outcome of many individual images. Similarly, a pair of scalp electrodes can not "see" individual brain potentials on the other side of relatively thick skull bone. What it does see is the momentary movement of thousands of signals that happen to be "marching" in the same direction, but this direction changes many times in one second.
- - -
12-3 Electroencephalographic (EEG) Signals
In 1924, Hans Berger connected two electrodes (small, round, metal disks) to a patient's scalp and detected a feeble current by using a very delicate galvanometer. Berger thereby opened up a window on the brain. As electronic amplifiers and recording equipment came into common use, the brain signal - called an electroencephalogram (EEG) - became a major clinical tool of the neurologist [J. D. Bronzino, 1984; R. Elul, 1972; A. S. Gevins, 1984; A. S. Gevins et al., 1975; K. A. Kooi, R. P. Tucker & R. E. Marshall, 1978; M. Matonsek et al., 1967; G. Pfurtscheller and R. Cooper, 1975; A. Remond, 1971; E. Vaadia, H. Bergman & M. Abeles, 1989].
Five EEG records taken from a normal individual are shown in Fig. 12-4 [H. H. Jasper, 1941]. The time scale is given by the 1-s line at the bottom. The voltage scales are given by the 50- mV 1 lines at the right side. For this type of recording, the "hot" electrode is usually located on the back of the head, opposite the striate cortex, and the other on a neutral point such as an ear lobe. In order to get a good recording, one must of course make good contact with the skin. It is not necessary to shave the patient's head, but skin oil should be removed with a solvent. A conducting paste is applied to the electrode, which is held by a suitable strap or cap against the scalp. The neutral ear-lobe electrode can be held in place with a simple clip.
[S.D. writes: "When I was at the Rutgers Medical School in New Jersey, I designed equipment for telemetering 15 simultaneous EEGs to a receiver in an adjoining room, via FM, so that the animal or human patient could be

Fig. 12-4 Electroencephalographic recordings taken from a normal individual. For this type of recording, the "hot" electrode is usually located on the back of the head, the other on a neutral point such as an ear lobe. In the "relaxed" waveform the patient's eyes are closed. Notice the time and voltage scales: the 1-s line at the bottom and 50-mV 1 lines at the right side. (From Jasper, Epilepsy and cerebral localization, Thomas, 1941.)
free to move about. In the first test of the equipment I volunteered to be the patient. My neurologist friend and colleague insisted that the normal solvent treatment was inadequate for my oily skin, and proceeded to sandpaper my ear lobe until it bled. The recording was normal, but I was the buff of jokes for the next week or two about how my ear became bitten. The moral of the story-don't let a Ph.D. connect EEG electrodes to a patient or anybody else."]
The ear-lobe electrode does not pick up appreciable signals because it is relatively far away from electrical sources such as brain cells and muscle fibers; in fact, that is why it is considered to be a neutral point. The other electrode only "sees" the electrical activity in its immediate vicinity. To begin with, because of the thickness of scalp, muscles, skull bone, and mem- brane surrounding the brain, the electrode is around 1 cm away from the cerebral cortex. Signals from neurons deeper in the brain are rapidly attenuated as distance increases. The traces of Fig. 12-4 therefore reveal local happenings. The voltage from an individual neuron is far too small to show up on an electrode l cm away. What we are seeing are the voltages from thousands of neurons whose action or graded potentials happen to be headed in the same direction at the same time. It is somewhat like being up in a satellite and trying to see individuals on earth. If everybody in Times Square suddenly headed north, say, you could possibly see the flow in one direction. Normally, however, people moving one way cancel out the people moving in the opposite direction.
In the first waveform of Fig. 12-4, the patient is "excited" (but not agitated) in the sense that he (or she) is looking at the people in the room and listening to their conversations. To avoid interference from the APs that elicit muscle contraction, the patient is not talking or moving about. We see that the EEG is a weak (10 mV 1 ), chaotic signal. From the frequencies present in the trace, we can conclude that it is not derived from high-speed (100 meters/second (m/s» APs, which would induce relatively high frequencies as they fly by. It is not even derived from slow 1 m/s APs. The frequencies are reminiscent of the 0.1-m/s progression of a graded potential of maximum amplitude 20 mV as it is conducted along a dendrite: its neuron integrates many inputs to generate its own graded potential, and the latter is "leisurely"
transmitted to another dendrite via an electrical gap junction or via a chemical agent that diffuses across a synaptic junction.
The fear voiced by some individuals that the neurologist can read their "inner thoughts" is without foundation. Certain regularities can be observed if the person is looking at, say, a pattern of vertical stripes, but the same thought repeated over and over again merely results in a random, nonrepetitive and meaningless EEG pattern [Skeptical Jnquirer].
In the second waveform, which is "relaxed," the patient' s eyes are closed. This state gives rise to a relatively strong, nonsteady oscillation at around 10 Hz, known as the alpha rhythm [H. T. Castello, 1983; B. H. Jansen, 1985; J. G. Okyere, P. Y. Ktonas & J. S. Meyer, 1986]. The origin of the alpha rhythm is controversial, but it is probably connected to vision because it is strongest in the vicinity of the striate cortex. Especially mysterious is the fact that it vanishes when the patient opens his or her eyes and looks at the world about. A hypothetical explanation is offered at the end of this section.
As the patient progresses (or retrogresses) from relaxed to drowsy to asleep to deep sleep, radical changes in frequency and amplitude take place. "Deep sleep" displays slow waves, only a few cycles per second, of large amplitude (100 mV 1). The large amplitude indicates that entire sections of tissue beneath the electrode are undergoing the same electrochemical changes in unison. The slow waves hint at the relatively slow transport of jons. Perhaps the EEG is telling us that the debris following many hours of wakefulness is being removed and "normal" material is being replenished. Altematively, in Chapter II, it is suggested that self-organization lakes place during "quiet" periods such as sleep. These are relatively slow processes; the human cycle consists of 16 hours of alertness followed by eight hours of sleep (not all of it, of course, as "deep sleep") [A. R. Morrison, 1983].
The clinical value of the EEG comes from the fact that an abnormal waveform is correlated with abnormalcy underneath the electrode [N. I. Bachen, 1986; D. G. Childers et al., 1982; C. D. McGillem, J. I. Aunon & K.-B. Vu, 1985; N. H. Morgan and A. S. Gevins, 1986; G. Pfurtscheller,
UN MÉTODO DE ESTIMULACIÓN POR CONTROL REMOTO DEL SISTEMA NERVIOSO
Esta información procede de la revista de Biología y Medicina de la universidad de Yale. Precisamente la universidad donde el Dr. Delgado estaba trabajando, antes de aceptar la dirección de un equipo de investigadores en la Universidad Autónoma de Madrid invitado por el Ministro del gobierno del Opus Dei de Franco, Villar Palasí, para investigar sobre estimulaciones cerebrales no invasivas (sin implantes).
From The Yale Journal of Biology and Medicine, Vol. 7, No. 1, October 1934, New Haven, Conn.
A METHOD FOR THE REMOTE CONTROL OF
ELECTRICAL STIMULATION OF THE
NERVOUS SYSTEM*
E. LEON CHAFFEE AND RICHARD U. LIGHT
I. The History of Electrical Excitation
In his studies of function the physiologist has been greatly dependent on the basic sciences, not only in the attempt to identify the separate processes, but also in the interpretation of their actions. Particularly has this been true of investigations of the neuromuscular system, where electrophysics has paved the way to many important discoveries. It may even be said that the history of neurophysiology has been decided in large part by the development of electric-recording instruments on the one hand, and by the increasingly effective use of electric currents for stimulating on the other. As Adrian has written: "It would be hard to think of any other method which has done so much to show us how the body works, for it gives us a means of throwing a muscle or a nerve into activity at will by an agency which does no damage and can be precisely controlled." 1
The early developments of these two sciences went forward hand in hand, since many of the discoveries in electricity were due to the tell-tale sensations and spasms caused by the passage of a current;in the absence of precise instruments for measuring electrical currents, the unique susceptibility of the neuromuscular system to electrical excitation made it an indispensable detector. Thus it was that in 1746 a group af scientists in Leyden encountered unexpectedly the "capacity" of an electrified glass of water, because of the sudden shock which accompanied its discharge. The sensation was described vividly by one of the group: "Mr. Muschenbroeck, who tried the experiment with a very thin glass bowl, says, in a letter to Mr. Réaumur, which he wrote soon after the experiment,
* From the Cruft Laboratory of Physics and Engineering, Harvard University, and the Department of Surgery, Yale University School of Medicine. A preliminary report of this method was published in Science, March 30, 1934, Vol. 79 pp. 299-300
YALE JOURNAL OF BIOLOGY AND MEDICINE, VOL. 7, No.2
--------------------------------------------------------------------------------
84 YALE JOURNAL OF BIOLOGY AND MEDICINE
that he felt himself struck in his arms, shoulders and breast, so that he lost his breath, and was two days before he recovered from the effects of the blow and the terror. He adds, that he would not take a second shock for the kingdom of France." 24 In such manner began the history of electrical excitation, together with the recognition of the first known reservoir of electrical energy, the famed Leyden jar.
Medicine, in its traditional weakness, promptly embraced the new spark phenomenon as a therapeutic agent, and applied it with little hesitancy to human ailments. During the years which followed the announcement of the "Leyden Phial" there was a wave of new cures. Most of these hinged on the fantastic belief that if odoriferous medicines were confined in glass vessels, and the vessels were excited electrically, the .medicinal virtues would transpire through the glass to be absorbed by the patients in whose hands the containers were placed! 25 The fallacy of these ill-judged methods was soon to be exposed by the Abbé Nollet, 20 but the procedure of electrification meanwhile had been transferred directly to the patient, :and numerous cures of paralyses were being recorded. Among the most optimistic of the "electrotherapists" was John Wesley the divine who, although possessing neither medical nor scientific degrees, yet organized dispensaries for the treatment of disease, and used "this unparalleled remedy" as one of his principal therapeutic agents. His book, 31 published in 1759, is replete with miracles which tested the patience of men of stricter scientific discipline. The remedy fell into better hands when Jean Paul Marat adopted it in his lucrative medical practice, but he too wrote with an enthusiasm which exceeded the evident results. 21 In the hands of charlatans it was an ideal tool, and the notorious Graham, 29 who exploited the English public unmercifully, was the first of a never-ending succes sion of quacks who have used the mysterious nature of electricity to further their deceptions.
The interest of Benjamin Franklin in electrotherapy appeared at an early date, for in the early fifties he had gathered a number of paralyzed patients from eastern Pennsylvania for systematic daily treatment in his laboratory, by application of a series of condenser discharges of fairly high capacity. His battery of jars yielded sparks great enough to produce local hyperthermia, petechiae, and paræsthesias. Occasionally the treatments resulted in some increase
--------------------------------------------------------------------------------
ELECTRICAL EXCITATION 85
of voluntary motion, and he wrote to a friend: "These appearances gave great spirits to the patients, and made them hope a perfect cure; but I do not remember, that I ever saw any amendment after the fifth day; which the patients perceiving, and finding the shocks pretty severe, they became discouraged, went home, and a short time relapsed; so that in palsies, I never knew any advantage from electricity, that was permanent. And how far the temporary advantage might arise from the exercise of the patient's journey, and coming daily to my house, or from the spirits, given by the hope of success, enabling them to exert more strength in moving their limbs, I shall not pretend to say." 10 Sagacious Franklin! He recognized the effectiveness of this measure in the treatment of psychiatric disorders, but he held little belief in the wildly hailed specificity of its action.
It was logical that electric sparks should one day be applied to exposed nerves and muscles, and to Galvani and his' coworkers belongs the credit for this crucial experiment, which resulted in the discovery of the electrical excitability of nerves (1780-83 ). 12 It was an important finding, for not only did it open up new methods in the study of neurology, but it provided a detector of electric currents far better than tactile sensation, and thus paved the way for Galvani's second great contribution, the discovery of chemically generated electric current (1789). 13 The quantity of electricity necessary to evoke a convulsion in the freshly killed frog's leg is exceedingly minute, and Volta 30 found it to be 50 to 60 times less than that which could be detected by the most sensitive electrometer of the day, so that for 30 years the nerve-muscle preparation (called by Volta "the animal electrometer") occupied a unique position in the physicist's laboratory.
The rapidly growing science of electricity was soon to demand instruments of greater pre'cision, however, and the epochal work of Oersted 23 (1820) in linking the two forces of galvanism and magnetism provided a basis for the detection of a current by its magnetic effect. The instrument constructed on this principle was called a "galvanometer", and the appellation is proper not only for its descriptive force, but because the device marked the first advance over Galvani's other "meter", the animal electrometer.
At this time Magendie (1822), while utilizing the galvanic current to supplement his studies of the spinal roots, demonstrated
--------------------------------------------------------------------------------
86 YALE JOURNAL OF BIOLOGY AND MEDICINE
that stimulation of the anterior roots causes muscular contraction and that of posterior roots sensation. 20 His studies were of fundamental importance to an understanding of the neurological structure, and they gave rise to a type of experiment which since has been steadily repeated: first, to reduce the activity of an organ through nerve section, and second, to increase its activity (or certain phases thereof) through nerve stimulation. This method has been at the root of much of our present knowledge of neurology, but it lacks a certain balance in that the results which follow division of a nerve are virtually permanent, whereas those attendant upon stimulation are of the briefest duration,-for the current can be applied only during the course of a surgical exposure.
It had not escaped the attention of Galvani that muscular contraction occurs at the beginning and at the end of current flow, while the muscle remains quiescent during the period of steady flow. This characteristic made possible a series of repeated stimuli from a source of interrupted current, and in 1845 DuBois-Reymond 6 began to apply the methods for generating alternating currents recently uncovered by Faraday to the uses of physiology. His device, an
induction coil with a magnetically driven breaker in the primary circuit, produced a series of closely spaced stimuli as the current went through its cycles. The frequency of the current pulses was varied in a rough way through adjustments of the spring tension, and the amount of current could be controlled by changes in the degree of magnetic coupling. This mechanism, simple and effective, is still to be found in every physiological laboratory today. It has,
however, three disadvantages: it operates at unknown frequencies, except when regulated by tuning forks; the precise current developed is unknown; and the wave-form used for stimulation is irregular and varies in shape from one coil to another and with varying conditions of use. The latter characteristic has come into much prominence of recent years, largely through the demonstrations by
Lapicque 17 and Keith Lucas' 19 of the quantity called "chronaxie", but even the careful studies of such men as Erlanger and Garrey 7 have not succeeded in stabilizing the output characteristics of the DuBois-Reymond (or "Harvard") coils. It is to be hoped, therefore, that the venerable induction coil will soon be supplanted by low-frequency oscillators employing thermionic vacuum tubes, so that investigations carried on in widely separated laboratories may
--------------------------------------------------------------------------------
ELECTRICAL EXCITATION 87
be reported in terms of the common units of electrophysics and thus be understandable to all.
Electrical stimulation of peripheral nerves has revealed the functions of many tissues, through the exaggerated activity of the parts to which they lead. However, nerves are merely conduction paths, and the controlling action is known to lie deeper, in the spinal cord or the brain. For many years, attempts had been made to elicit responses from the application of currents directly to the surface of the brain, but these resulted in failure until Fritsch and Hitzig 11 (1870) announced that electrical excitation of certain small areas of the cerebral cortex of the dog would give rise to muscular movements in the opposite side of the body. Much interest attaches to this finding, as it has led to the modern views of localization of cerebral function in the hands of Ferrier, 8 Beevor and Horsley, 2 Sherrington, 27 and the host of modern investigators, who have added definition to the picture of cerebral activity by the use of this method. The studies have not been confined to animals alone, however, for as early as 1909 Cushing 5 pointed to the results of electrical stimulation of the postcentral gyrus during the course of surgical removal of intracranial tumors.
As soon as one considers the use of stimulating currents in physiological investigations, he must choose between the employment of two closely spaced electrodes (bipolar) and the method of grounding the body to a large indifferent plate, leaving only one wire as an active electrode (monopolar). The relative efficacy of the two systems has been the cause of much controversy, although so far as the brain is concerned, it appears that properly constructed electrodes are as satisfactory in pairs as in a grounded circuit. 28 In peripheral nerve studies, however, an important distinction exists, for it must not be forgotten that Galvani's original experiments were made with monopolar stimulation; and that Volta's successful argument against "animal electricity" hinged on the application of both electrodes to the nerve, thus showing that a nervous impulse and not a conducted current traversed the remaining nerve trunk to set the muscle into contraction.
Until very recent years physiologists have been content to observe the effects produced by direct electrical excitation during the short period of a crucial experiment in the decorticate animal, or in one under general anæsthesia. With the growing recognition of the
--------------------------------------------------------------------------------
88 YALE JOURNAL OF BIOLOGY AND MEDICINE
immense duties of the central nervous system in the regulation of autonomic functions, it has become apparent that no method of study can succeed in revealing these central controlling mechanisms, that does not permit the student to reproduce artificially a controlling force which is similar to the function under investigation. This conception is best examined by a comparative consideration of the motor apparatus, for in this system the activities are directive, precise, and capable of attaining full accomplishment in a few moments. They respond to individual electric stimuli, even under anæsthesia, and the shocks may be given with fair rapidity since the cycle of activity can be completed many times in the course of a second. Cerebral autonomic centers, on the other hand, are invested with the regulation of the ceaseless mechanism of internallife, in which changes are of degree and not of the fundamental act, and ebb and flow succeed each other at a comparatively leisurely pace. The very existence of higher autonomic centers has only been suggested, largely as the result of accident or disease, and they have proved extraordinarily refractory to experimental demonstration. It is quite possible that the difficulty rests with the investigative method employed, for not only does it lack the continuing character of automatism, but it is usually applied together with a general anæsthesia, the depressing action of which is exerted on the very region under consideration.
Several efforts have been made to simulate more nearly the functional character of these processes, as well as to reach the other neurological systems during their normal phases of activity. In 1915 Keeton and Becht 15 during an experimental study or the pituitary body in dogs, implanted some iron filings directly into the gland. The dogs were allowed to recover, and were then brought within the range of an electromagnet in the hope that local stimulation might occur at the site of the implanted iron. The experiment failed to demonstrate any resulting change and this was attributed to the weak magnetic fields available 16 although in any case there would have resulted only a movement of the filings (mechanical stimulation) and not an induced electric current. In Zürich, Hess 14 has developed a technic for bringing conducting wires out through the skin to form a direct electrical circuit, and his patience and thoroughness in handling this difficult method are an inspiration to those who visit his laboratory. In this country Mussen 22 Bradford Cannon, 3 and others have adopted similar means. These workers
--------------------------------------------------------------------------------
ELECTRICAL EXCITATION 89
have succeeded in prolonging the duration of the experiment from a few hours at the time of the operation, to a period of several days after recovery. The drawback to the method lies in the wires and clamps used, which project through the skin, and are therefore susceptible to accident and facilitate infection, so that it is seldom that the experiment can be continued longer than two or three weeks. Furthermore, the arrangement requires the constant watch and restraint of the animal during the time that wires are connected to the source of current, so that the subject does not enjoy real freedom, nor can the experiment be adjusted to the changing phases of physiology.
The germ of a new bio-electric relationship seems to have been demonstrated by Feser, 9 who reports the successful transmission of radio waves to an exposed nerve-muscle preparation, without the use of any receiving "pick-up" mechanism other than the nerve itself, which he believes acts as a dipole antenna. His experiments, which deserve more thorough substantiation, may prove useful to analytical studies of the nervous impulse.
The development toward which all of these experiments point is the complete separation of the animal from restraint. The complications of the sterile surgical field, and the time limit set by the operative procedure, must be avoided. Anæsthesia, which suppresses many functions, should not be used during the investigation of the nervous system, yet the animal must not be led into awareness of the experiment through pain or restraint. Under the best conditions, the animal would be allowed to lead a normal life throughout the duration of the experiment, without interference of eating and drinking, sleeping, or exercising. The actual excitation should be confined to the small area of the nervous system under consideration and be applied without rear or current spreading. Since the degree or stimulation depends in part upon the current density in the excited tissue, and also on the manner in which the current varies with time4 these factors must be under ready control or the operator. Because, too, the character or the response is orten dependent on the number or times per minute or per second that the stimulus is repeated, provision must be made ror accurate and rapid adjustments of the rate or stimulation.
With these difficult requirements in mind, the junior author enlisted the aid or Proressor Chaffee in designing a circuit which would be simple, effective, and durable and which would incorporate precision methods of electrophysics, so that subsequent physiological analysis might rest on ground as secure as possible.
Professor Chaffee has taken keen interest in the problem, and gives in the next section his development of the circuits employed.*
*The apparatus requires the use of a small secondary coil, implanted in the animal during a preliminary surgical operation. It is of interest that during the past year two other groups have also come independently to the use of buried collodion-coated coils for this purpose- Loucks, 18 in Baltimore, during an attempt to test the effect of repeted cerebral stimuli in creating conditioned reflexes, implanted small coils beneath the cervical skin of dogs, which received current through inductive coupling to a primary coil tied to the dog's neck. Fender and Scott (Rochester, New York) have informed us of tests coducted with an apparatus in which a primary coil of several hundred turns surrounds a small box, while the terminals of the imbedded secondary coil are led to a vasomotor nerve.
II Design and operation of the apparatus.
In order to produce electrical stimulation a current of electricity is caused to flow through the tissue to be excited, the degree of stimulation being dependent upon the maximum current density and the way the current varies with time. The exciting current is caused to flow between two electrodes applied to the tissue by connecting the electrodes to some source of electrical energy, such as a battery, an induction coil, or a source of alternating current. If the nerve or muscle being excited is excised or is a part of anaesthetized animal, the connecting wires from the preparation to the electrical source cause little or no interference or inconvenience. If, however, it is desired to study the effect of stimulating definite portions of the brain or nervous system of an animal in its normal state and perhaps over extended periods of time, connecting wires cannot be employed. The method to be described permits stimulation through an electromagnetic connection instead of by a physical connection, and hence eliminates the undesirable features of previous methods.
An important feature of the method to be described in this paper is that fairly exact knowledge of and control of the intensity and wave form of the stimulating current are possible.
Principle of the Method. The bare principle of operation of the method is briefly as follows: A large condenser, charged from a source of direct current, is periodically discharged through a few turns of primary coil three or more feet in diameter. This discharge is very intense, amounting to about a thousand amperes, although of short duration.The resulting intense magnetic field produced by this discharge induces an electromotive force in a sterile secondary coil of many turns of fine wire implanted beneath the skin of the animal located withon the large primary coil. One terminal of the secondary coil is connected to a silver plate imbedded anywhere in the moist tissue of the animal. This electrode is inert with respect to stimulation effects. The other terminal of the coil is led by a fine insulated wire to a spot to be stimulated. Only a small area approximately equal to the cross section of this wire makes

[C2 100 pF
R2' 10000 ohms
C2' 0.05 uf
L2 2000 turns 0.17 H // 700 ohms
E0 5 kW at 500V
R0=50-30000 carries 10 A and dissipates all the power]
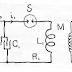
Fig 1. Schematic Circuit Diagram.
contact with the tissue. The potential induced in the secondary coil causes current impulses to flow between the two electrodes. The current density is maximum at the exposed surface of the fine wire electrode and hence, if the current is not too great, stimulation is confined to a very small volume of tissue at the end of the stimulating electrode.
The two coils described sbove are actually the primary and secondary coils of an induction coil, the secondary coil being entirely embedded inside the animal. In order, however, that sufficient potentials be produced in the secondary coil the electrical elements must have quite different characteristics from those of the ordinary induction coil.
A simplified wiring diagram of the essential elements of the system is shown in Fig 1 Letters L1 and R1 indicate the inductance and resistance of the large primary coil.*
* In the mathematical analysis to follow L1 and R1 stand for the inductance and resistance of the entire primary circuit.
C1 is a large condenser charged from a source of power of voltage E0 through the variable resistance R0. The condenser C1 is discharged periodically through L1 by means of a contacting device S which closes and opens the circuit at an adjustable frequency. The secondary coil has inductance indicated by L2 and resistance indicated by R2. The coil and wires have a certain distributed capacitance represented by the ditted condenser C2, The conduction path through the tissue being stimulated has a resistance R2', and an equivalent series capacitance C2' which is due to the electrolytic polarization at the terminals
- - -
[skipping the rest of p 92]
- - -
--------------------------------------------------------------------------------
ELECTRICAL EXCITATION 93
The resistance of the primary circuit varies with the number of turns in the primary coil and also with the frequency because of increasing skin effect with increasing frequency. The resistance of the entire primary circuit, exclusive of the interrupter S, was measured by means of a resonance bridge at the resonance frequency of the circuit. The results are given in Table II:
TABLE II
RESISTANCE AND FREQUENCY OF PRIMARY CIRCUIT
C, = 73.8mf
Primary coil 36 inches diameter.
Frequency in cycles Resistance
Turns per second of circuit
Turn 2
Frequency in cycles per second 5030
Resistance of circuit 0.011 ohms
Turn 3
Frequency in cycles per second 3740
Resistance of circuit 0.015 ohms
Turn 4
Frequency in cycles per second 2950
Resistance of circuit 0.021 ohms
The strength of field varies with position inside the primary coil, but as shown later the variation is most rapid near the periphery. The variation of field was found not to be excessive within a cubic cage 20 inches on a side placed within a coil 36 inches in diameter. Monkeys and small baboons can live comfortably in such a cage. With this experimental arrangement voltages as high as 110 volts could be obtained in the secondary coil, which indicate that a considerably larger primary coil can to advantage be used, permitting either a larger cage or a smaller variation of field within the same cage. The interrupter S was first a mechanically operated contact between large silver-tipped terminals. Sparking rapidly spoils such a contact. For this reason and also to eliminate the noise of the contactor, a special mercury-pool vacuum tube was used with circuits to be described later. This tube, type FG 158, is a new development of the General Electric Company and at the request of the company details of construction cannot be given at this time. The characteristics of the tube are as follows: When the tube is not conducting its resistance is very great. Conduction through the tube is initiated by impressing a high potential from an induction coil
--------------------------------------------------------------------------------
94 YALE JOURNAL OF BIOLOGY AND MEDICINE
between the mercury pool and an outside band of metal surrounding the tube just above the mercury. The resistance of the tube rapidly drops to a very low value as the current attains values of the order of a thousand amperes. The resistance of the tube varies with the current but the voltage drop across the tube is approximately constant at about 14 volts during conduction.
Resistance R, is variable from a minimum of about 50 ohms to a maximum of about 30,000 ohms. This resistance is made up of several units, and each unit must have adequate current-carrying capacity because practically all of the power supplied by the generator Eo is lost in this resistance. The low values of resistance can carry up to 10 amperes and dissipate five kilowatts of power. The source of power Eo is capable of delivering up to 5 kilowatts at 500 volts.
The design of the secondary coil to be implanted in the subject depends upon many factors. The physical dimensions are limited on the one hand by the size and shape that can conveniently be implanted, while on the other hand the magnitude of electrical potential produced for stimulation depends in part upon the area of the coil and the cross section of the winding space. The coil adopted for implanting in a circular hole cut in the skull of a small monkey has an outside diameter of 1.5 " and a thickness of 0.125" with a winding depth in a radial direction of 0.25". Such a coil is shown in Fig 2.
The number of turns of wire with which the secondary coil should be wound to give the best results is best determined by mathematical analysis to be given later, and depends upon the frequency used and the resistance R'2 of the tissue between the terminals. The standard coil adopted for most of the experiments has 2000 turns of No. 40 (B & S gauge) enamelled copper wire. Such a coil has an inductance of 0.17 henries and a resistance of 700 ohms. For the same winding space the inductance and resistance are practically proportional to the square of the number of turns.

Fig 2. Secondary Coil.
[Repeating some data from the text below
silver plate 0.5", platinum wire 2.5" coated along its length appart from the electrode tip having a crossection of 0.0000497 inch2= 0.000321 cm2, primary discharge:frequencies 2.4 -5 kHz, rep rate up to 14 Hz linear and above that chaotic. The maximum value of the current density at the stimulating electrode is the peak potential divided by R2' and by the crossectional area of the wire electrode. For example if the peak potential is 10V and R2' is 10000 ohms the peak current density is 3.12 A/cm2]
- - -
--------------------------------------------------------------------------------
ELECTRICAL EXCITATION 95
The ends of the coil are soldered to short lengths of No. 32 platinum wire; the joint and inner ends of the platinum wire are securely bound to the coil with silk thread; and then the coil is impregnated and coated with collodion. The platinum-wire leads previously enamelled are also covered with a fairly thick insulating coat of collodion. One platinum wire is cut to a length of approximately 1¼ inches and soldered to a plate of silver ½ inch square. The other platinum-wire terminal is approximately 2½ inches long and is coated throughout its length, the only area exposed being the cross section of the wire at the perpendicularly cut end. This area is 49.7 X 10-6 sq. inches, or 3.21 X 10-4 cm2. The distributed capacitance of the secondary coil in air is approximately 12 micro-micro-farads (mmf), but when the coil and its wire terminals are immersed in a 1/10 normal KCI solution to simulate the tissue in which the coil is implanted, the distributed capacitance represented by C2 of Fig. 1 is about 100mmf. Each inch added to the length of the pair of terminal wires adds approximately 5mmf to the distributed capacitance. The electrical path through the tissue between the terminals of the secondary coil possesses resistance and capacitance. The capacitance is partly polarization capacitance at the interface between the tissue and the stimulating electrode, and undoubtedly partly capacitance due to the cell walls in the tissue. The capacitance and resistance vary with the potential applied across the tissue path and to some extent with the frequency. At a single frequency we can approximately represent the electrical characteristics of the tissue by a resistance R'2 in series with a capacitance C'2 as indicated in Fig. 1. A number of measurements were made by means of a bridge of the resistance R'2 and the capacitance C'2 between copper wires of various sizes and a plate, the wires and plate being imbedded in various locations in the brain of an anæsthetized monkey. The values of R'2 and C'2 found varied considerably for the same size wire, due probably to varying amounts of moisture at the end of the wire electrode. Average values for R'2 and C'2 for a No. 32 B & S wire are 10,000 ohms and 0.05 mf. If R'2 is larger C'2 is less, their product being approximately constant. Since a knowledge of the values of these two quantities is essential to determining the intensity of stimulation, a method will be given later of measuring R'2 and C'2 in situ.
- - -
[Skipping p 96-118. On p 118 -119 the Triple discharge circuit in fig 15 is discussed
A single circuit produces impulses varying widely depending on the orientation. The Triple Discharge Circuit is a generalized method that uses three perpendicular circuits excited in such a way that the stimulating current varies between 0.7 and 1 of maximum and the wireless connection to the experimental animal becomes correspondingly more stable]
--------------------------------------------------------------------------------
118

FIG. 13. Three Primary Coils.

FIG. 14. Cage Inside Primary Coils.
--------------------------------------------------------------------------------
119


FIG. 16. X-ray film of a coil implanted in a young baboon. The coil occupies a space in the left occipital region from which a circular area of bone has been removed. The silver plate lies behind the scalp, and the platinum wire terminal, insulated except at its tip, is resting in the hand area of the right motor cortex.
--------------------------------------------------------------------------------
FIG. 15. Triple Circuit Diagram .

--------------------------------------------------------------------------------
120
- - -
III. EXPERIENCES WITH THE APPARATUS IN USE
The examples which follow indicate something of the practical use of the method, although many of the tests were made early in its development. Experiments with motor and sensory nerves gave evidence that the mechanism was operating as planned, and also afforded an excellent check on the surgical technic. A cerebral implantation was then carried out with the coil inserted in place of a
ELECTRICAL EXCITATION 121
circular area of cranium, as shown in Fig. 16, and the active terminal was laid on the motor cortex. Again the result was definite, with the appearance of an unexpected relationship between the frequency of impulses and the character of the response, to be described in the third protocol. One example is given of implantation in a "silent" center, and another shows the effects of vagal excitation, but thus far attention has been directed chiefly toward the development of a system which would prove easy to control in the situations in which the result was predictable.
Experiences with these animals have made it plain that the experimenter must be able not only to control the various factors of his circuit, but must have accurate indicators for these during the progress of the experiment. It follows from Professor Chaffee's description that the duration of the stimulus depends upon the resonance frequency of the primary circuit, a factor which can be found either by calculation or by bridge measurements. The frequency of repetition of the stimulus can now be measured directly on an ammeter and the cumbersome stroboscopic unit has been discarded except for calibration of the ammeter dial. The voltage of stimulation from the buried coil is measured by feeding the current pickup from a duplicate coil into a vacuum-tube voltmeter, but the determination is complicated somewhat by the changes of resistance between the buried electrodes, which must first be determined on an impedance bridge. From a knowledge of these elements the current density can be computed.
In spite of the fact that the central autonomic system was our first goal, the apparatus cannot be considered to have anyone special field of usefulness. The dramatic duplication of the picture of epilepsy during remotely controlled stimulation of the cerebral cortex has thrown emphasis to the possibilities of restudying other neurological functions. Most localization studies have been carried out under the handicaps of the older methods, and it is probable that not only in the case of the motor cortex, but in the spinal cord and peripheral nerves as well, new characteristics may be brought out by properly regulated stimulation given during the course of the ordinary day's living.
Protocols
Excitation of a pure motor nerve (hypoglossal) in a dog by induction of current into a secondary coil implanted in the neck.
On November 3,
--------------------------------------------------------------------------------
122 YALE JOURNAL OF BIOLOGY AND MEDICINE
1933, a medium-sized dog was operated on under nembutal anæsthesia and the right hypoglossal nerve was exposed and stimulated for identification with the ordinary close-coupled inductor. A small secondary coil was inserted into a recess in the subcutaneous tissue and the platinum electrode was brought to rest just within the nerve sheath. The indifferent electrode, which consisted of a silver plate ½ inch square, was laid beneath the subcutaneous tissue.
The coil and its wires were fastened in place by silk sutures and the wound was closed. Before outlining the results obtained with this implanted coil, it is worth while to describe the apparatus in use at the time. The primary coil consisted of two turns and its diameter was one foot. The discharge mechanism was operated by a small electric motor driving through two cams. When one of these raised a tappet the line current (220 v.) was connected with the condensers (80 mf.). After the condensers were charged the charging current was disconnected by opening this contact, and the second cam came into position and joined the capacitors with the primary coil through which the discharge then occurred. With this arrangement (and three pairs of inter changeable gears interposed between the electric motor and the cam assembly) it was possible to vary the frequency of discharge from individual impulses spaced three or four seconds apart, up to a frequency of about 20 discharges per second. At these higher speeds the mechanism was noisy and much heat was generated at the contacts, although these were constructed of ½ inch silver discs. Continued operation of 10 or I5 minutes at this speed resulted in the disintegration of the contacts and it was required that they be renewed during the experiment. A commutating mechanism of this type was therefore wholly inadequate for voltages which were used subsequently (450 v.) and for operating speeds of 50-100 per second, and the mechanism was discarded in favor of a vacuum-tube design.
However difficult was the manipulation and maintenance of this apparatus, it still served to demonstrate that the principle could be successfully applied. Stimulation experiments were carried out in this animal both at the conclusion of the operation (with anæsthesia still present) and on the following day. The results differed little, except for the opportunity for observing the animal's own reaction to the unexpected muscular movements of the tongue. The primary coil was brought to a position parallel with the implanted coil and separated from it by about 12 inches. It was found that the right half of the tongue responded to individual discharges as slowly as they might be given. As the frequency of discharge* was increased the muscle still gave individual contractions for each impulse given, until the rate
* "Frequency of discharge" refers to the number of times per second that the condensers are charged and discharged;-not to the self-tuned frequency of the circuit, which is of the order of 3000 cycles.
--------------------------------------------------------------------------------
ELECTRICAL EXCITATION 123
reached 15 per second. Above this rate the response appeared to be tetanic in nature. During the tests made on the day after operation, the animal stood quietly and was not disturbed by the experience. He made no attempt to draw away from the primary coil and was not alarmed by the unusual motion of the tongue.
2. Stimulation of the vagus nerve (thoracic portion) of a dog with simultaneous collection of the gastric juice. On November 9, 1933, an operation was performed under sodium amytal anæsthesia on a dog of medium size to expose the lower portion of the esophagus through a transthoracic approach. The intercostal muscles were divided between the 6th and 7th ribs in the right axillary line and the ribs were separated. The pleura was opened and lobes of the right lung held back with gauze packs. The vagus nerves were identified on the lower esophagus, where they consist of anterior and posterior trunks. A small secondary coil was implanted outside the chest wall just posterior to the incision and the indifferent electrode was placed in the subcutaneous tissue. The active terminal was brought between the ribs and made to lie along the posterior border of the thorax where its tip was sutured so as to be in contact with the posterior branch of the vagus nerve after a small incision was made in the nerve sheath. The electrode lay about 1 ½ inches above the diaphragm. Careful closure was carried out.
On the following day the dog was awake and moving around actively. The wound was dry, and the buried coil there was not evident except on palpation through the skin. A stomach tube was passed and a fasting specimen of gastric juice was obtained. This was of small amount (2.5 cc.) and showed no free acid but contained a total acidity of 70°. Stimulation was then begun (10: 15 A. M.) at a rate of 15 impulses per second for an uninterrupted period of 5 minutes. No abdominal or diaphragmatic muscle spasm was seen and the dog apparently was not aware of the act of stimulation. Two minutes after the stimulation was started gastric juice began to flow from the tube and for fifteen minutes the flow continued intermittently, without the necessity for application of negative pressure to the tube. Stimulation was halted at 10:20 and at 10:30 forty cubic centimeters of light-brown, watery gastric juice had been collected, which was found to be devoid of free acid, but with total acidity of 120°. At 10:50 a third specimen was withdrawn (26 cc.) without free acid, and with 115° total acid. A second period of stimulation was carried out from 10: 50 to 10: 55 at a rate of 18 impulses per second. On this occasion it was noted that violent peristalsis began two minutes after stimulation was started and continued for 20 minutes. This was so active that not only could it be heard through the stethoscope, but it was visible over the entire surface of the abdomen. No bowel movements occurred, nor was there diarrhea following the experiment. At 11:05, 21 cc. of gastric juice had been collected and showed total acidity of 115°.
--------------------------------------------------------------------------------
124 YALE JOURNAL OF BIOLOGY AND MEDICINE
The last specimen was taken at 11:20 and amounted to 13 cc. with free acid of 10° and total acid 105°.
It was recognized at the time of operation that the respiratory movements of the esophagus and diaphragm were of such magnitude that the tiny wire which was sutured to the vagus nerve on the esophagus would not remain in place long without being broken, and an attempt at stimulation on the fifth postoperative day bore out this prediction. The use of fine wire-stranded cable has been suggested by F. A. Fender, to permit mobility without breakage, but it is probably true that an experiment of long duration can be carried out best with this system when the coil and its terminals are implanted in rigid locations such as the skull, the posterior portion of the thorax, and the pelvis. In the case of the vagus, the nerve should be partially transplanted to a retropleural position. This form of study of the action of the vagus and the splanchnic nerves should yield important information concerning the nature of digestion, for it thus provides the means whereby stimulation can be applied in the conscious vegetating animal during the actual time that the various digestive phases are under way. It should also prove of value in identifying the effects of long-continued excessive vagus and splanchnic activity in relation to the problem of peptic ulcer, and perhaps to other disease processes within the abdomen.
3. Stimulation of the motor cortex of a Mocacus rhesus. On April 27, 1934, operation was performed on an immature monkey under sodium amytal anæsthesia. A left fronto-parietal flap was lifted back, and a coil was implanted in a second opening made by removing a circular area of bone in the right occipital region. The broad plate terminal was laid outside the dura, and the left motor cortex was explored with the active electrode. The operative procedure had been simplified by constructing a special wooden operating table on the under surface of which was fastened the primary coil of the discharge mechanism. The magnetic field extended well above the level of the animal's head, so that with the mechanism in operation currents were generated in the small coil without the necessity of bringing wires into the sterile field. The platinum terminal was laid in the hand region of area 4. Two weeks later recovery Was complete, and the first tests were made. The machine was set to give impulses at 14 per second. So soon as the circuit was thrown into operation, the animal's right arm went into adduction and extension, and the reaction was so pronounced that the current was stopped after only two seconds of stimulation had been received. The right arm continued spastic, and developed a tremor which spread in Jacksonian style to
--------------------------------------------------------------------------------
ELECTRICAL EXCITATION 125
involve the entire right side of the body. The seizure lasted nearly a minute, but recovery was fairly prompt. After a short rest, stimulation was repeated at the same rate, for a period of 30 seconds. The response in the right hand began immediately, and within 3 or 4 seconds the entire right side was in clonic motion. Very quickly he collapsed to the floor of the cage, and a rhythmical motion (3 or 4 twitches per second) appeared in both arms and both legs. To all appearances he was unconscious, and was breathing noisily, while the face bore a deep cyanotic appearance. In his convulsive motions he struck the back of the head against the cage, and drew a little blood. The seizure subsided gradually over a period of two minutes; he lay quietly for a moment, and then awakened and rose. He was very unsteady on his feet for 15 or 20 seconds and fell down several times, looking as if bewildered. A minute later he was completely normal again. Although it is impossible here to give a full description of the experiences with stimulation of the motor cortex in conscious animals, it is worth noting that the rate of stimulation affects the type of response. So long as the rate is kept below 12 or 13 impulses per second, the response from area 4 consists of discrete muscular contractions which appear coincident with the firing of the tube. When the rate of discharge rises above this level, the response affects more distant areas, and may be either a unilateral or a completely bilateral seizure. Furthermore, the rhythm of shaking becomes about 3 movements per second, instead of small fibrillary twitches in time with the succession of electrical stimuli. This sharp change in response, which occurs while the voltage, current density, and duration of stimulus are kept constant, but when the rate of stimulation is increased, suggests strongly the importance of investigating the counteractive effects of drugs commonly used to reduce or prevent epileptic attacks, while maintaining careful control of the electrical constants of the circuit.
4. Investigation of the left premotor area of cortex in a Macacus rhesus (3.5 kg.). The coil was implanted in this animal on May 7, 1934, under light ether anæsthesia. A left boneflap was made, as well as a circular craniotomy on the right for the coil, and the premotor area was identified with a platinum electrode before finally suturing it in place. When the stimulus was applied to area 4 (Brodmann's classification), the response was immediate, in the form of jerky motions of the opposite side synchronous with the current surges from the condensers. As the electrode was moved forward into area 6 the response in the opposite arm became more delayed, like a slowly twisting
--------------------------------------------------------------------------------
126 YALE JOURNAL OF BIOLOGY AND MEDICINE
cramp, with a distinct tendency to continue after the current had ceased. The terminal was placed upon the premotor hand area, a centimeter rostral to the edge of area 4.
The animal recovered from the operation without incident, and on May 24th was first studied with the stimulation device, before a gathering of the Halsted Club. The reactions are described in the words of Dr. William Mahoney, who conducted the experiment: The animal was placed within the magnetic field and a rate of stimulation of 12 impulses per second was given. Almost immediately there began a series of chewing and tonguewagging motions. Nothing else happened for a time, but after a minute or perhaps longer the right hand began to assume the position shown in such pictures as are usually presented in text-books for "cerebellar hand," that is, extension of the hand combined with flexion at the wrist. Shortly afterward there appeared clonic motions of the arm, and the animal whirled about on its ischial protuberances and fell to its right side. The clonic jerks then extended into the right leg. The stimulus was stopped after about two minutes, but the seizure persisted for a minute longer, and involved the left side of the body as well as the right side. Then for a time he lay on the left side with the left arm and leg extended, and the right arm and leg flexed. There was no deviation of the eyes, nor anisocoria, and adversive movements of the head were not noticed. Gradually he relaxed, and arose to a sitting posture, but he appeared dazed. The face was flushed, and for a few minutes the right hand hung limply as he moved about. He had voided at the beginning of stimulation.
5. Experiments with a monkey in which an electrode was inserted in the hypothalamus. On Nov. 17, 1933, an adult male Macacus rhesus weighing 6.5 kg. was operated on under sodium amytal anæsthesia, and a coil was placed in a circular opening in the calvarium. The indifferent electrode was laid beneath the scalp, and the active platinum electrode was brought forward over the surface of the frontal lobe, which was elevated to permit insertion of the wires into the tissue just anterior to the optic chiasm. The tip of the wire was pushed backwards about 1 cm. from this point into the hypothalamus.
Ten days later wound healing was complete, and the animal was stimulated for a period of ten minutes at 10 impulses per second. At first he was restless and moved about the cage, then he became quiet, and yawned several times. There was no urination, pupillary change, nor bowel movements. He remained quiet until the end of the period. Two minutes after stimulation ceased the animal became drowsy, and his head drooped. It was difficult to arouse him with a stick, although five minutes later he woke up for a short time and took interest in his surroundings. His head was soon down again, and for 5 minutes he appeared to be asleep. He was finally aroused by a noise, but not until 30 minutes after the stimulation did he become bright and active.
--------------------------------------------------------------------------------
ELECTRICAL EXCITATION 127
On a second period of stimulation the same results followed, but we noticed also that there was a general vasodilatation over the entire skin which faded out as the current was stopped, and started when it was recommenced.
In summary a method is presented for the study of excitable regions in the nervous system during the normal life of an animal. It employs two coils which are actually the primary and secondary elements of an induction coil. The secondary coil is entirely imbedded inside the animal and the primary coil is of large size and carries currents of high peak value. The animal is permitted to live comfortably within a cage through which passes the magnetic field from the primary coil. Stimulation occurs at one of the terminals of the secondary coil which can be placed in any portion of the central or peripheral nervous system. A description is given of the electrical circuits employed and several examples are cited to indicate the range of usefulness of the apparatus.
BIBLIOGRAPHY
1) Adrian, E. D.: The Mechanism of Nervous Action. Philadelphia, Univ. of Penn. Press, 1932, p. 3.
2) Beevor, A. and Horsley, V.: A minute analysis (experimental) of the various movements produced by stimulating in the monkey different regions of the cortical center for the upper limb, as defined by Professor Ferrier. Phil. Trans. Roy. Soc., London, 1887,178B, 153-67.
3) Cannon, B.: A melhod of stimulating autonomic nerves in the unanæsthetized cat with observations on the motor and sensory effects. Am. J. Physiol., 1933, 105, 366-72.
4) Cole, K. S.: Electric excitation in nerve. Cold Spring Harbor Symposia on Quantitative Biology, 1933, 1,1-13.
5) Cushing, H.: Faradic stimulation of postcentral gyrus in conscious patients. Brain, 1909, 32, 44-53.
6) DuBois-Reymond, E.: Untersuchungen über thierische Elektricität. Berlin, 1848.
7) Erlanger, J. and Garrey, W. E.: Faradic stimuli: A physical and physiological study. Am. J. Physiol., 1914, 35, 377-473.
8) Ferrier, D.: The Functions of the Brain. G. P. Putnam's Sons, N. Y., 1880.
9) Feser, J.: Drahtlose Reizung der Froschnerven mittels elektrischer Wellen. Deut. tierärtz. Wchschr., 1930, 38, 23-4.
10) Franklin, B., in a Jetter to Dr. Pringle, read before the Royal Society, Jan. 12, 1758. Quoted in part in (2), pp. 412-13. Reprinted in full in The American Museum: or Repository of Ancient and Modern Fugitive Pieces, etc., Prose and Poetical. 1789, 6, 53-4.
11) Fritsch, G. and Hitzig, E.: Ueber die elektrische Erregbarkeit des Grosshims. Archiv. f. Anat., Physiol. u. wissen. Med., Leipzig, 1870, p. 300.
--------------------------------------------------------------------------------
128 YALE JOURNAL OF BIOLOGY AND MEDICINE
12) Galvani, Luigi: Collezione delle Opera, Bologna, 1841, pp. 17-27. [See also:] Dalton, J. C.: The experimental Method in Medical Science. Lecture 1. Galvani and Galvanism in the Study of the Nervous System. N. Y., G. P. Putnam's Sons, 1882.
13) Galvani, Luigi: De viribus electricitatis in motu musculari, Commentarius. Bologna, 1791.
14) Hess, W. R.: Beiträge zur Physiologie des Hirnstammes. 1. Teil. Die Methodik der lokalisierten Reizung und Ausschaltung subkortikaler Hin abschnitte. Leipzig, 1932.
15) Keeton, R. W. and Becht, F. C.: The stimulation of the hypophysis in dogs. Amer. J. Physiol., 1915, 39, 109-22.
16) Keeton, R. W.: Personal communication to Dr. William Mahoney, April, 1934.
17) Lapicque, L., and Lapicque, Mme. L.: Sur la forme de la loi d'excitation electrique exprimee par la quantite. Reponse a M. Hoorweg. Compt. rend. Soc. de biol., 1905, 58, 668-70.
18) Loucks, R. B.: Preliminary report of a technique for stimulation or destruction of tissues beneath the integument and the establishing of conditioned reactions with faradization of the cerebral cortex. J. Comp. Psychol., 1933, 16, 439-44.
19) Lucas, K.: The conduction of the nervous impulse. London, 1917.
20) Magendie, F.: Experiences sur les fonctions des racines des nerfs rachidiens. J. de Physiol. Exper. et Path., 1822, t. ii, p. 276.
21) Marat, J. P.: Memoire sur l'electricite medicale. Paris, Méquignon, 1784.
22) Mussen, A. T.: Cerebellum and Red Nucleus: A preliminary report on a new method of physiologic investigation. Arch. Neurol. & Psychiat., 1934, 31, 1 10-26.
23) Oersted, H. C.: Experimenta circa effectum conflictus electrici in acum magneticam. Copenhagen, 1820.
24) Priestley, J.: The History and Present State of Electricity, with Original Experiments. London, 1767, p. 83.
25) Ibid., p. 146-7.
26) Ibid., p. 152.
27) Sherrington, C. S.: The integrative action of the nervous system. New Haven, Yale U niv. Press, 1906.
28) Smith, W. K.: Motor cortex of the bear (Ursus americanus). Arch. Neurol. & Psychiat., 1933,30,14-39.
29) Turrell, W. J.: Three Electrotherapists of the Eighteenth Century. Ann. Med. Hist., 1921,3,361-67.
30) Volta, A.: An account of some discoveries made by M. Galvani, with experiments and observations on them. London, Phil. Trans. Roy. Soc., 1793.
31) Wesley, J.: The Desideratum: or, Electricity made Plain and Useful. By a Lover of Mankind and of Common Sense. London, 1759.
From The Yale Journal of Biology and Medicine, Vol. 7, No. 1, October 1934, New Haven, Conn.
A METHOD FOR THE REMOTE CONTROL OF
ELECTRICAL STIMULATION OF THE
NERVOUS SYSTEM*
E. LEON CHAFFEE AND RICHARD U. LIGHT
I. The History of Electrical Excitation
In his studies of function the physiologist has been greatly dependent on the basic sciences, not only in the attempt to identify the separate processes, but also in the interpretation of their actions. Particularly has this been true of investigations of the neuromuscular system, where electrophysics has paved the way to many important discoveries. It may even be said that the history of neurophysiology has been decided in large part by the development of electric-recording instruments on the one hand, and by the increasingly effective use of electric currents for stimulating on the other. As Adrian has written: "It would be hard to think of any other method which has done so much to show us how the body works, for it gives us a means of throwing a muscle or a nerve into activity at will by an agency which does no damage and can be precisely controlled." 1
The early developments of these two sciences went forward hand in hand, since many of the discoveries in electricity were due to the tell-tale sensations and spasms caused by the passage of a current;in the absence of precise instruments for measuring electrical currents, the unique susceptibility of the neuromuscular system to electrical excitation made it an indispensable detector. Thus it was that in 1746 a group af scientists in Leyden encountered unexpectedly the "capacity" of an electrified glass of water, because of the sudden shock which accompanied its discharge. The sensation was described vividly by one of the group: "Mr. Muschenbroeck, who tried the experiment with a very thin glass bowl, says, in a letter to Mr. Réaumur, which he wrote soon after the experiment,
* From the Cruft Laboratory of Physics and Engineering, Harvard University, and the Department of Surgery, Yale University School of Medicine. A preliminary report of this method was published in Science, March 30, 1934, Vol. 79 pp. 299-300
YALE JOURNAL OF BIOLOGY AND MEDICINE, VOL. 7, No.2
--------------------------------------------------------------------------------
84 YALE JOURNAL OF BIOLOGY AND MEDICINE
that he felt himself struck in his arms, shoulders and breast, so that he lost his breath, and was two days before he recovered from the effects of the blow and the terror. He adds, that he would not take a second shock for the kingdom of France." 24 In such manner began the history of electrical excitation, together with the recognition of the first known reservoir of electrical energy, the famed Leyden jar.
Medicine, in its traditional weakness, promptly embraced the new spark phenomenon as a therapeutic agent, and applied it with little hesitancy to human ailments. During the years which followed the announcement of the "Leyden Phial" there was a wave of new cures. Most of these hinged on the fantastic belief that if odoriferous medicines were confined in glass vessels, and the vessels were excited electrically, the .medicinal virtues would transpire through the glass to be absorbed by the patients in whose hands the containers were placed! 25 The fallacy of these ill-judged methods was soon to be exposed by the Abbé Nollet, 20 but the procedure of electrification meanwhile had been transferred directly to the patient, :and numerous cures of paralyses were being recorded. Among the most optimistic of the "electrotherapists" was John Wesley the divine who, although possessing neither medical nor scientific degrees, yet organized dispensaries for the treatment of disease, and used "this unparalleled remedy" as one of his principal therapeutic agents. His book, 31 published in 1759, is replete with miracles which tested the patience of men of stricter scientific discipline. The remedy fell into better hands when Jean Paul Marat adopted it in his lucrative medical practice, but he too wrote with an enthusiasm which exceeded the evident results. 21 In the hands of charlatans it was an ideal tool, and the notorious Graham, 29 who exploited the English public unmercifully, was the first of a never-ending succes sion of quacks who have used the mysterious nature of electricity to further their deceptions.
The interest of Benjamin Franklin in electrotherapy appeared at an early date, for in the early fifties he had gathered a number of paralyzed patients from eastern Pennsylvania for systematic daily treatment in his laboratory, by application of a series of condenser discharges of fairly high capacity. His battery of jars yielded sparks great enough to produce local hyperthermia, petechiae, and paræsthesias. Occasionally the treatments resulted in some increase
--------------------------------------------------------------------------------
ELECTRICAL EXCITATION 85
of voluntary motion, and he wrote to a friend: "These appearances gave great spirits to the patients, and made them hope a perfect cure; but I do not remember, that I ever saw any amendment after the fifth day; which the patients perceiving, and finding the shocks pretty severe, they became discouraged, went home, and a short time relapsed; so that in palsies, I never knew any advantage from electricity, that was permanent. And how far the temporary advantage might arise from the exercise of the patient's journey, and coming daily to my house, or from the spirits, given by the hope of success, enabling them to exert more strength in moving their limbs, I shall not pretend to say." 10 Sagacious Franklin! He recognized the effectiveness of this measure in the treatment of psychiatric disorders, but he held little belief in the wildly hailed specificity of its action.
It was logical that electric sparks should one day be applied to exposed nerves and muscles, and to Galvani and his' coworkers belongs the credit for this crucial experiment, which resulted in the discovery of the electrical excitability of nerves (1780-83 ). 12 It was an important finding, for not only did it open up new methods in the study of neurology, but it provided a detector of electric currents far better than tactile sensation, and thus paved the way for Galvani's second great contribution, the discovery of chemically generated electric current (1789). 13 The quantity of electricity necessary to evoke a convulsion in the freshly killed frog's leg is exceedingly minute, and Volta 30 found it to be 50 to 60 times less than that which could be detected by the most sensitive electrometer of the day, so that for 30 years the nerve-muscle preparation (called by Volta "the animal electrometer") occupied a unique position in the physicist's laboratory.
The rapidly growing science of electricity was soon to demand instruments of greater pre'cision, however, and the epochal work of Oersted 23 (1820) in linking the two forces of galvanism and magnetism provided a basis for the detection of a current by its magnetic effect. The instrument constructed on this principle was called a "galvanometer", and the appellation is proper not only for its descriptive force, but because the device marked the first advance over Galvani's other "meter", the animal electrometer.
At this time Magendie (1822), while utilizing the galvanic current to supplement his studies of the spinal roots, demonstrated
--------------------------------------------------------------------------------
86 YALE JOURNAL OF BIOLOGY AND MEDICINE
that stimulation of the anterior roots causes muscular contraction and that of posterior roots sensation. 20 His studies were of fundamental importance to an understanding of the neurological structure, and they gave rise to a type of experiment which since has been steadily repeated: first, to reduce the activity of an organ through nerve section, and second, to increase its activity (or certain phases thereof) through nerve stimulation. This method has been at the root of much of our present knowledge of neurology, but it lacks a certain balance in that the results which follow division of a nerve are virtually permanent, whereas those attendant upon stimulation are of the briefest duration,-for the current can be applied only during the course of a surgical exposure.
It had not escaped the attention of Galvani that muscular contraction occurs at the beginning and at the end of current flow, while the muscle remains quiescent during the period of steady flow. This characteristic made possible a series of repeated stimuli from a source of interrupted current, and in 1845 DuBois-Reymond 6 began to apply the methods for generating alternating currents recently uncovered by Faraday to the uses of physiology. His device, an
induction coil with a magnetically driven breaker in the primary circuit, produced a series of closely spaced stimuli as the current went through its cycles. The frequency of the current pulses was varied in a rough way through adjustments of the spring tension, and the amount of current could be controlled by changes in the degree of magnetic coupling. This mechanism, simple and effective, is still to be found in every physiological laboratory today. It has,
however, three disadvantages: it operates at unknown frequencies, except when regulated by tuning forks; the precise current developed is unknown; and the wave-form used for stimulation is irregular and varies in shape from one coil to another and with varying conditions of use. The latter characteristic has come into much prominence of recent years, largely through the demonstrations by
Lapicque 17 and Keith Lucas' 19 of the quantity called "chronaxie", but even the careful studies of such men as Erlanger and Garrey 7 have not succeeded in stabilizing the output characteristics of the DuBois-Reymond (or "Harvard") coils. It is to be hoped, therefore, that the venerable induction coil will soon be supplanted by low-frequency oscillators employing thermionic vacuum tubes, so that investigations carried on in widely separated laboratories may
--------------------------------------------------------------------------------
ELECTRICAL EXCITATION 87
be reported in terms of the common units of electrophysics and thus be understandable to all.
Electrical stimulation of peripheral nerves has revealed the functions of many tissues, through the exaggerated activity of the parts to which they lead. However, nerves are merely conduction paths, and the controlling action is known to lie deeper, in the spinal cord or the brain. For many years, attempts had been made to elicit responses from the application of currents directly to the surface of the brain, but these resulted in failure until Fritsch and Hitzig 11 (1870) announced that electrical excitation of certain small areas of the cerebral cortex of the dog would give rise to muscular movements in the opposite side of the body. Much interest attaches to this finding, as it has led to the modern views of localization of cerebral function in the hands of Ferrier, 8 Beevor and Horsley, 2 Sherrington, 27 and the host of modern investigators, who have added definition to the picture of cerebral activity by the use of this method. The studies have not been confined to animals alone, however, for as early as 1909 Cushing 5 pointed to the results of electrical stimulation of the postcentral gyrus during the course of surgical removal of intracranial tumors.
As soon as one considers the use of stimulating currents in physiological investigations, he must choose between the employment of two closely spaced electrodes (bipolar) and the method of grounding the body to a large indifferent plate, leaving only one wire as an active electrode (monopolar). The relative efficacy of the two systems has been the cause of much controversy, although so far as the brain is concerned, it appears that properly constructed electrodes are as satisfactory in pairs as in a grounded circuit. 28 In peripheral nerve studies, however, an important distinction exists, for it must not be forgotten that Galvani's original experiments were made with monopolar stimulation; and that Volta's successful argument against "animal electricity" hinged on the application of both electrodes to the nerve, thus showing that a nervous impulse and not a conducted current traversed the remaining nerve trunk to set the muscle into contraction.
Until very recent years physiologists have been content to observe the effects produced by direct electrical excitation during the short period of a crucial experiment in the decorticate animal, or in one under general anæsthesia. With the growing recognition of the
--------------------------------------------------------------------------------
88 YALE JOURNAL OF BIOLOGY AND MEDICINE
immense duties of the central nervous system in the regulation of autonomic functions, it has become apparent that no method of study can succeed in revealing these central controlling mechanisms, that does not permit the student to reproduce artificially a controlling force which is similar to the function under investigation. This conception is best examined by a comparative consideration of the motor apparatus, for in this system the activities are directive, precise, and capable of attaining full accomplishment in a few moments. They respond to individual electric stimuli, even under anæsthesia, and the shocks may be given with fair rapidity since the cycle of activity can be completed many times in the course of a second. Cerebral autonomic centers, on the other hand, are invested with the regulation of the ceaseless mechanism of internallife, in which changes are of degree and not of the fundamental act, and ebb and flow succeed each other at a comparatively leisurely pace. The very existence of higher autonomic centers has only been suggested, largely as the result of accident or disease, and they have proved extraordinarily refractory to experimental demonstration. It is quite possible that the difficulty rests with the investigative method employed, for not only does it lack the continuing character of automatism, but it is usually applied together with a general anæsthesia, the depressing action of which is exerted on the very region under consideration.
Several efforts have been made to simulate more nearly the functional character of these processes, as well as to reach the other neurological systems during their normal phases of activity. In 1915 Keeton and Becht 15 during an experimental study or the pituitary body in dogs, implanted some iron filings directly into the gland. The dogs were allowed to recover, and were then brought within the range of an electromagnet in the hope that local stimulation might occur at the site of the implanted iron. The experiment failed to demonstrate any resulting change and this was attributed to the weak magnetic fields available 16 although in any case there would have resulted only a movement of the filings (mechanical stimulation) and not an induced electric current. In Zürich, Hess 14 has developed a technic for bringing conducting wires out through the skin to form a direct electrical circuit, and his patience and thoroughness in handling this difficult method are an inspiration to those who visit his laboratory. In this country Mussen 22 Bradford Cannon, 3 and others have adopted similar means. These workers
--------------------------------------------------------------------------------
ELECTRICAL EXCITATION 89
have succeeded in prolonging the duration of the experiment from a few hours at the time of the operation, to a period of several days after recovery. The drawback to the method lies in the wires and clamps used, which project through the skin, and are therefore susceptible to accident and facilitate infection, so that it is seldom that the experiment can be continued longer than two or three weeks. Furthermore, the arrangement requires the constant watch and restraint of the animal during the time that wires are connected to the source of current, so that the subject does not enjoy real freedom, nor can the experiment be adjusted to the changing phases of physiology.
The germ of a new bio-electric relationship seems to have been demonstrated by Feser, 9 who reports the successful transmission of radio waves to an exposed nerve-muscle preparation, without the use of any receiving "pick-up" mechanism other than the nerve itself, which he believes acts as a dipole antenna. His experiments, which deserve more thorough substantiation, may prove useful to analytical studies of the nervous impulse.
The development toward which all of these experiments point is the complete separation of the animal from restraint. The complications of the sterile surgical field, and the time limit set by the operative procedure, must be avoided. Anæsthesia, which suppresses many functions, should not be used during the investigation of the nervous system, yet the animal must not be led into awareness of the experiment through pain or restraint. Under the best conditions, the animal would be allowed to lead a normal life throughout the duration of the experiment, without interference of eating and drinking, sleeping, or exercising. The actual excitation should be confined to the small area of the nervous system under consideration and be applied without rear or current spreading. Since the degree or stimulation depends in part upon the current density in the excited tissue, and also on the manner in which the current varies with time4 these factors must be under ready control or the operator. Because, too, the character or the response is orten dependent on the number or times per minute or per second that the stimulus is repeated, provision must be made ror accurate and rapid adjustments of the rate or stimulation.
With these difficult requirements in mind, the junior author enlisted the aid or Proressor Chaffee in designing a circuit which would be simple, effective, and durable and which would incorporate precision methods of electrophysics, so that subsequent physiological analysis might rest on ground as secure as possible.
Professor Chaffee has taken keen interest in the problem, and gives in the next section his development of the circuits employed.*
*The apparatus requires the use of a small secondary coil, implanted in the animal during a preliminary surgical operation. It is of interest that during the past year two other groups have also come independently to the use of buried collodion-coated coils for this purpose- Loucks, 18 in Baltimore, during an attempt to test the effect of repeted cerebral stimuli in creating conditioned reflexes, implanted small coils beneath the cervical skin of dogs, which received current through inductive coupling to a primary coil tied to the dog's neck. Fender and Scott (Rochester, New York) have informed us of tests coducted with an apparatus in which a primary coil of several hundred turns surrounds a small box, while the terminals of the imbedded secondary coil are led to a vasomotor nerve.
II Design and operation of the apparatus.
In order to produce electrical stimulation a current of electricity is caused to flow through the tissue to be excited, the degree of stimulation being dependent upon the maximum current density and the way the current varies with time. The exciting current is caused to flow between two electrodes applied to the tissue by connecting the electrodes to some source of electrical energy, such as a battery, an induction coil, or a source of alternating current. If the nerve or muscle being excited is excised or is a part of anaesthetized animal, the connecting wires from the preparation to the electrical source cause little or no interference or inconvenience. If, however, it is desired to study the effect of stimulating definite portions of the brain or nervous system of an animal in its normal state and perhaps over extended periods of time, connecting wires cannot be employed. The method to be described permits stimulation through an electromagnetic connection instead of by a physical connection, and hence eliminates the undesirable features of previous methods.
An important feature of the method to be described in this paper is that fairly exact knowledge of and control of the intensity and wave form of the stimulating current are possible.
Principle of the Method. The bare principle of operation of the method is briefly as follows: A large condenser, charged from a source of direct current, is periodically discharged through a few turns of primary coil three or more feet in diameter. This discharge is very intense, amounting to about a thousand amperes, although of short duration.The resulting intense magnetic field produced by this discharge induces an electromotive force in a sterile secondary coil of many turns of fine wire implanted beneath the skin of the animal located withon the large primary coil. One terminal of the secondary coil is connected to a silver plate imbedded anywhere in the moist tissue of the animal. This electrode is inert with respect to stimulation effects. The other terminal of the coil is led by a fine insulated wire to a spot to be stimulated. Only a small area approximately equal to the cross section of this wire makes

[C2 100 pF
R2' 10000 ohms
C2' 0.05 uf
L2 2000 turns 0.17 H // 700 ohms
E0 5 kW at 500V
R0=50-30000 carries 10 A and dissipates all the power]
Fig 1. Schematic Circuit Diagram.
contact with the tissue. The potential induced in the secondary coil causes current impulses to flow between the two electrodes. The current density is maximum at the exposed surface of the fine wire electrode and hence, if the current is not too great, stimulation is confined to a very small volume of tissue at the end of the stimulating electrode.
The two coils described sbove are actually the primary and secondary coils of an induction coil, the secondary coil being entirely embedded inside the animal. In order, however, that sufficient potentials be produced in the secondary coil the electrical elements must have quite different characteristics from those of the ordinary induction coil.
A simplified wiring diagram of the essential elements of the system is shown in Fig 1 Letters L1 and R1 indicate the inductance and resistance of the large primary coil.*
* In the mathematical analysis to follow L1 and R1 stand for the inductance and resistance of the entire primary circuit.
C1 is a large condenser charged from a source of power of voltage E0 through the variable resistance R0. The condenser C1 is discharged periodically through L1 by means of a contacting device S which closes and opens the circuit at an adjustable frequency. The secondary coil has inductance indicated by L2 and resistance indicated by R2. The coil and wires have a certain distributed capacitance represented by the ditted condenser C2, The conduction path through the tissue being stimulated has a resistance R2', and an equivalent series capacitance C2' which is due to the electrolytic polarization at the terminals
- - -
[skipping the rest of p 92]
- - -
--------------------------------------------------------------------------------
ELECTRICAL EXCITATION 93
The resistance of the primary circuit varies with the number of turns in the primary coil and also with the frequency because of increasing skin effect with increasing frequency. The resistance of the entire primary circuit, exclusive of the interrupter S, was measured by means of a resonance bridge at the resonance frequency of the circuit. The results are given in Table II:
TABLE II
RESISTANCE AND FREQUENCY OF PRIMARY CIRCUIT
C, = 73.8mf
Primary coil 36 inches diameter.
Frequency in cycles Resistance
Turns per second of circuit
Turn 2
Frequency in cycles per second 5030
Resistance of circuit 0.011 ohms
Turn 3
Frequency in cycles per second 3740
Resistance of circuit 0.015 ohms
Turn 4
Frequency in cycles per second 2950
Resistance of circuit 0.021 ohms
The strength of field varies with position inside the primary coil, but as shown later the variation is most rapid near the periphery. The variation of field was found not to be excessive within a cubic cage 20 inches on a side placed within a coil 36 inches in diameter. Monkeys and small baboons can live comfortably in such a cage. With this experimental arrangement voltages as high as 110 volts could be obtained in the secondary coil, which indicate that a considerably larger primary coil can to advantage be used, permitting either a larger cage or a smaller variation of field within the same cage. The interrupter S was first a mechanically operated contact between large silver-tipped terminals. Sparking rapidly spoils such a contact. For this reason and also to eliminate the noise of the contactor, a special mercury-pool vacuum tube was used with circuits to be described later. This tube, type FG 158, is a new development of the General Electric Company and at the request of the company details of construction cannot be given at this time. The characteristics of the tube are as follows: When the tube is not conducting its resistance is very great. Conduction through the tube is initiated by impressing a high potential from an induction coil
--------------------------------------------------------------------------------
94 YALE JOURNAL OF BIOLOGY AND MEDICINE
between the mercury pool and an outside band of metal surrounding the tube just above the mercury. The resistance of the tube rapidly drops to a very low value as the current attains values of the order of a thousand amperes. The resistance of the tube varies with the current but the voltage drop across the tube is approximately constant at about 14 volts during conduction.
Resistance R, is variable from a minimum of about 50 ohms to a maximum of about 30,000 ohms. This resistance is made up of several units, and each unit must have adequate current-carrying capacity because practically all of the power supplied by the generator Eo is lost in this resistance. The low values of resistance can carry up to 10 amperes and dissipate five kilowatts of power. The source of power Eo is capable of delivering up to 5 kilowatts at 500 volts.
The design of the secondary coil to be implanted in the subject depends upon many factors. The physical dimensions are limited on the one hand by the size and shape that can conveniently be implanted, while on the other hand the magnitude of electrical potential produced for stimulation depends in part upon the area of the coil and the cross section of the winding space. The coil adopted for implanting in a circular hole cut in the skull of a small monkey has an outside diameter of 1.5 " and a thickness of 0.125" with a winding depth in a radial direction of 0.25". Such a coil is shown in Fig 2.
The number of turns of wire with which the secondary coil should be wound to give the best results is best determined by mathematical analysis to be given later, and depends upon the frequency used and the resistance R'2 of the tissue between the terminals. The standard coil adopted for most of the experiments has 2000 turns of No. 40 (B & S gauge) enamelled copper wire. Such a coil has an inductance of 0.17 henries and a resistance of 700 ohms. For the same winding space the inductance and resistance are practically proportional to the square of the number of turns.

Fig 2. Secondary Coil.
[Repeating some data from the text below
silver plate 0.5", platinum wire 2.5" coated along its length appart from the electrode tip having a crossection of 0.0000497 inch2= 0.000321 cm2, primary discharge:frequencies 2.4 -5 kHz, rep rate up to 14 Hz linear and above that chaotic. The maximum value of the current density at the stimulating electrode is the peak potential divided by R2' and by the crossectional area of the wire electrode. For example if the peak potential is 10V and R2' is 10000 ohms the peak current density is 3.12 A/cm2]
- - -
--------------------------------------------------------------------------------
ELECTRICAL EXCITATION 95
The ends of the coil are soldered to short lengths of No. 32 platinum wire; the joint and inner ends of the platinum wire are securely bound to the coil with silk thread; and then the coil is impregnated and coated with collodion. The platinum-wire leads previously enamelled are also covered with a fairly thick insulating coat of collodion. One platinum wire is cut to a length of approximately 1¼ inches and soldered to a plate of silver ½ inch square. The other platinum-wire terminal is approximately 2½ inches long and is coated throughout its length, the only area exposed being the cross section of the wire at the perpendicularly cut end. This area is 49.7 X 10-6 sq. inches, or 3.21 X 10-4 cm2. The distributed capacitance of the secondary coil in air is approximately 12 micro-micro-farads (mmf), but when the coil and its wire terminals are immersed in a 1/10 normal KCI solution to simulate the tissue in which the coil is implanted, the distributed capacitance represented by C2 of Fig. 1 is about 100mmf. Each inch added to the length of the pair of terminal wires adds approximately 5mmf to the distributed capacitance. The electrical path through the tissue between the terminals of the secondary coil possesses resistance and capacitance. The capacitance is partly polarization capacitance at the interface between the tissue and the stimulating electrode, and undoubtedly partly capacitance due to the cell walls in the tissue. The capacitance and resistance vary with the potential applied across the tissue path and to some extent with the frequency. At a single frequency we can approximately represent the electrical characteristics of the tissue by a resistance R'2 in series with a capacitance C'2 as indicated in Fig. 1. A number of measurements were made by means of a bridge of the resistance R'2 and the capacitance C'2 between copper wires of various sizes and a plate, the wires and plate being imbedded in various locations in the brain of an anæsthetized monkey. The values of R'2 and C'2 found varied considerably for the same size wire, due probably to varying amounts of moisture at the end of the wire electrode. Average values for R'2 and C'2 for a No. 32 B & S wire are 10,000 ohms and 0.05 mf. If R'2 is larger C'2 is less, their product being approximately constant. Since a knowledge of the values of these two quantities is essential to determining the intensity of stimulation, a method will be given later of measuring R'2 and C'2 in situ.
- - -
[Skipping p 96-118. On p 118 -119 the Triple discharge circuit in fig 15 is discussed
A single circuit produces impulses varying widely depending on the orientation. The Triple Discharge Circuit is a generalized method that uses three perpendicular circuits excited in such a way that the stimulating current varies between 0.7 and 1 of maximum and the wireless connection to the experimental animal becomes correspondingly more stable]
--------------------------------------------------------------------------------
118

FIG. 13. Three Primary Coils.

FIG. 14. Cage Inside Primary Coils.
--------------------------------------------------------------------------------
119


FIG. 16. X-ray film of a coil implanted in a young baboon. The coil occupies a space in the left occipital region from which a circular area of bone has been removed. The silver plate lies behind the scalp, and the platinum wire terminal, insulated except at its tip, is resting in the hand area of the right motor cortex.
--------------------------------------------------------------------------------
FIG. 15. Triple Circuit Diagram .

--------------------------------------------------------------------------------
120
- - -
III. EXPERIENCES WITH THE APPARATUS IN USE
The examples which follow indicate something of the practical use of the method, although many of the tests were made early in its development. Experiments with motor and sensory nerves gave evidence that the mechanism was operating as planned, and also afforded an excellent check on the surgical technic. A cerebral implantation was then carried out with the coil inserted in place of a
ELECTRICAL EXCITATION 121
circular area of cranium, as shown in Fig. 16, and the active terminal was laid on the motor cortex. Again the result was definite, with the appearance of an unexpected relationship between the frequency of impulses and the character of the response, to be described in the third protocol. One example is given of implantation in a "silent" center, and another shows the effects of vagal excitation, but thus far attention has been directed chiefly toward the development of a system which would prove easy to control in the situations in which the result was predictable.
Experiences with these animals have made it plain that the experimenter must be able not only to control the various factors of his circuit, but must have accurate indicators for these during the progress of the experiment. It follows from Professor Chaffee's description that the duration of the stimulus depends upon the resonance frequency of the primary circuit, a factor which can be found either by calculation or by bridge measurements. The frequency of repetition of the stimulus can now be measured directly on an ammeter and the cumbersome stroboscopic unit has been discarded except for calibration of the ammeter dial. The voltage of stimulation from the buried coil is measured by feeding the current pickup from a duplicate coil into a vacuum-tube voltmeter, but the determination is complicated somewhat by the changes of resistance between the buried electrodes, which must first be determined on an impedance bridge. From a knowledge of these elements the current density can be computed.
In spite of the fact that the central autonomic system was our first goal, the apparatus cannot be considered to have anyone special field of usefulness. The dramatic duplication of the picture of epilepsy during remotely controlled stimulation of the cerebral cortex has thrown emphasis to the possibilities of restudying other neurological functions. Most localization studies have been carried out under the handicaps of the older methods, and it is probable that not only in the case of the motor cortex, but in the spinal cord and peripheral nerves as well, new characteristics may be brought out by properly regulated stimulation given during the course of the ordinary day's living.
Protocols
Excitation of a pure motor nerve (hypoglossal) in a dog by induction of current into a secondary coil implanted in the neck.
On November 3,
--------------------------------------------------------------------------------
122 YALE JOURNAL OF BIOLOGY AND MEDICINE
1933, a medium-sized dog was operated on under nembutal anæsthesia and the right hypoglossal nerve was exposed and stimulated for identification with the ordinary close-coupled inductor. A small secondary coil was inserted into a recess in the subcutaneous tissue and the platinum electrode was brought to rest just within the nerve sheath. The indifferent electrode, which consisted of a silver plate ½ inch square, was laid beneath the subcutaneous tissue.
The coil and its wires were fastened in place by silk sutures and the wound was closed. Before outlining the results obtained with this implanted coil, it is worth while to describe the apparatus in use at the time. The primary coil consisted of two turns and its diameter was one foot. The discharge mechanism was operated by a small electric motor driving through two cams. When one of these raised a tappet the line current (220 v.) was connected with the condensers (80 mf.). After the condensers were charged the charging current was disconnected by opening this contact, and the second cam came into position and joined the capacitors with the primary coil through which the discharge then occurred. With this arrangement (and three pairs of inter changeable gears interposed between the electric motor and the cam assembly) it was possible to vary the frequency of discharge from individual impulses spaced three or four seconds apart, up to a frequency of about 20 discharges per second. At these higher speeds the mechanism was noisy and much heat was generated at the contacts, although these were constructed of ½ inch silver discs. Continued operation of 10 or I5 minutes at this speed resulted in the disintegration of the contacts and it was required that they be renewed during the experiment. A commutating mechanism of this type was therefore wholly inadequate for voltages which were used subsequently (450 v.) and for operating speeds of 50-100 per second, and the mechanism was discarded in favor of a vacuum-tube design.
However difficult was the manipulation and maintenance of this apparatus, it still served to demonstrate that the principle could be successfully applied. Stimulation experiments were carried out in this animal both at the conclusion of the operation (with anæsthesia still present) and on the following day. The results differed little, except for the opportunity for observing the animal's own reaction to the unexpected muscular movements of the tongue. The primary coil was brought to a position parallel with the implanted coil and separated from it by about 12 inches. It was found that the right half of the tongue responded to individual discharges as slowly as they might be given. As the frequency of discharge* was increased the muscle still gave individual contractions for each impulse given, until the rate
* "Frequency of discharge" refers to the number of times per second that the condensers are charged and discharged;-not to the self-tuned frequency of the circuit, which is of the order of 3000 cycles.
--------------------------------------------------------------------------------
ELECTRICAL EXCITATION 123
reached 15 per second. Above this rate the response appeared to be tetanic in nature. During the tests made on the day after operation, the animal stood quietly and was not disturbed by the experience. He made no attempt to draw away from the primary coil and was not alarmed by the unusual motion of the tongue.
2. Stimulation of the vagus nerve (thoracic portion) of a dog with simultaneous collection of the gastric juice. On November 9, 1933, an operation was performed under sodium amytal anæsthesia on a dog of medium size to expose the lower portion of the esophagus through a transthoracic approach. The intercostal muscles were divided between the 6th and 7th ribs in the right axillary line and the ribs were separated. The pleura was opened and lobes of the right lung held back with gauze packs. The vagus nerves were identified on the lower esophagus, where they consist of anterior and posterior trunks. A small secondary coil was implanted outside the chest wall just posterior to the incision and the indifferent electrode was placed in the subcutaneous tissue. The active terminal was brought between the ribs and made to lie along the posterior border of the thorax where its tip was sutured so as to be in contact with the posterior branch of the vagus nerve after a small incision was made in the nerve sheath. The electrode lay about 1 ½ inches above the diaphragm. Careful closure was carried out.
On the following day the dog was awake and moving around actively. The wound was dry, and the buried coil there was not evident except on palpation through the skin. A stomach tube was passed and a fasting specimen of gastric juice was obtained. This was of small amount (2.5 cc.) and showed no free acid but contained a total acidity of 70°. Stimulation was then begun (10: 15 A. M.) at a rate of 15 impulses per second for an uninterrupted period of 5 minutes. No abdominal or diaphragmatic muscle spasm was seen and the dog apparently was not aware of the act of stimulation. Two minutes after the stimulation was started gastric juice began to flow from the tube and for fifteen minutes the flow continued intermittently, without the necessity for application of negative pressure to the tube. Stimulation was halted at 10:20 and at 10:30 forty cubic centimeters of light-brown, watery gastric juice had been collected, which was found to be devoid of free acid, but with total acidity of 120°. At 10:50 a third specimen was withdrawn (26 cc.) without free acid, and with 115° total acid. A second period of stimulation was carried out from 10: 50 to 10: 55 at a rate of 18 impulses per second. On this occasion it was noted that violent peristalsis began two minutes after stimulation was started and continued for 20 minutes. This was so active that not only could it be heard through the stethoscope, but it was visible over the entire surface of the abdomen. No bowel movements occurred, nor was there diarrhea following the experiment. At 11:05, 21 cc. of gastric juice had been collected and showed total acidity of 115°.
--------------------------------------------------------------------------------
124 YALE JOURNAL OF BIOLOGY AND MEDICINE
The last specimen was taken at 11:20 and amounted to 13 cc. with free acid of 10° and total acid 105°.
It was recognized at the time of operation that the respiratory movements of the esophagus and diaphragm were of such magnitude that the tiny wire which was sutured to the vagus nerve on the esophagus would not remain in place long without being broken, and an attempt at stimulation on the fifth postoperative day bore out this prediction. The use of fine wire-stranded cable has been suggested by F. A. Fender, to permit mobility without breakage, but it is probably true that an experiment of long duration can be carried out best with this system when the coil and its terminals are implanted in rigid locations such as the skull, the posterior portion of the thorax, and the pelvis. In the case of the vagus, the nerve should be partially transplanted to a retropleural position. This form of study of the action of the vagus and the splanchnic nerves should yield important information concerning the nature of digestion, for it thus provides the means whereby stimulation can be applied in the conscious vegetating animal during the actual time that the various digestive phases are under way. It should also prove of value in identifying the effects of long-continued excessive vagus and splanchnic activity in relation to the problem of peptic ulcer, and perhaps to other disease processes within the abdomen.
3. Stimulation of the motor cortex of a Mocacus rhesus. On April 27, 1934, operation was performed on an immature monkey under sodium amytal anæsthesia. A left fronto-parietal flap was lifted back, and a coil was implanted in a second opening made by removing a circular area of bone in the right occipital region. The broad plate terminal was laid outside the dura, and the left motor cortex was explored with the active electrode. The operative procedure had been simplified by constructing a special wooden operating table on the under surface of which was fastened the primary coil of the discharge mechanism. The magnetic field extended well above the level of the animal's head, so that with the mechanism in operation currents were generated in the small coil without the necessity of bringing wires into the sterile field. The platinum terminal was laid in the hand region of area 4. Two weeks later recovery Was complete, and the first tests were made. The machine was set to give impulses at 14 per second. So soon as the circuit was thrown into operation, the animal's right arm went into adduction and extension, and the reaction was so pronounced that the current was stopped after only two seconds of stimulation had been received. The right arm continued spastic, and developed a tremor which spread in Jacksonian style to
--------------------------------------------------------------------------------
ELECTRICAL EXCITATION 125
involve the entire right side of the body. The seizure lasted nearly a minute, but recovery was fairly prompt. After a short rest, stimulation was repeated at the same rate, for a period of 30 seconds. The response in the right hand began immediately, and within 3 or 4 seconds the entire right side was in clonic motion. Very quickly he collapsed to the floor of the cage, and a rhythmical motion (3 or 4 twitches per second) appeared in both arms and both legs. To all appearances he was unconscious, and was breathing noisily, while the face bore a deep cyanotic appearance. In his convulsive motions he struck the back of the head against the cage, and drew a little blood. The seizure subsided gradually over a period of two minutes; he lay quietly for a moment, and then awakened and rose. He was very unsteady on his feet for 15 or 20 seconds and fell down several times, looking as if bewildered. A minute later he was completely normal again. Although it is impossible here to give a full description of the experiences with stimulation of the motor cortex in conscious animals, it is worth noting that the rate of stimulation affects the type of response. So long as the rate is kept below 12 or 13 impulses per second, the response from area 4 consists of discrete muscular contractions which appear coincident with the firing of the tube. When the rate of discharge rises above this level, the response affects more distant areas, and may be either a unilateral or a completely bilateral seizure. Furthermore, the rhythm of shaking becomes about 3 movements per second, instead of small fibrillary twitches in time with the succession of electrical stimuli. This sharp change in response, which occurs while the voltage, current density, and duration of stimulus are kept constant, but when the rate of stimulation is increased, suggests strongly the importance of investigating the counteractive effects of drugs commonly used to reduce or prevent epileptic attacks, while maintaining careful control of the electrical constants of the circuit.
4. Investigation of the left premotor area of cortex in a Macacus rhesus (3.5 kg.). The coil was implanted in this animal on May 7, 1934, under light ether anæsthesia. A left boneflap was made, as well as a circular craniotomy on the right for the coil, and the premotor area was identified with a platinum electrode before finally suturing it in place. When the stimulus was applied to area 4 (Brodmann's classification), the response was immediate, in the form of jerky motions of the opposite side synchronous with the current surges from the condensers. As the electrode was moved forward into area 6 the response in the opposite arm became more delayed, like a slowly twisting
--------------------------------------------------------------------------------
126 YALE JOURNAL OF BIOLOGY AND MEDICINE
cramp, with a distinct tendency to continue after the current had ceased. The terminal was placed upon the premotor hand area, a centimeter rostral to the edge of area 4.
The animal recovered from the operation without incident, and on May 24th was first studied with the stimulation device, before a gathering of the Halsted Club. The reactions are described in the words of Dr. William Mahoney, who conducted the experiment: The animal was placed within the magnetic field and a rate of stimulation of 12 impulses per second was given. Almost immediately there began a series of chewing and tonguewagging motions. Nothing else happened for a time, but after a minute or perhaps longer the right hand began to assume the position shown in such pictures as are usually presented in text-books for "cerebellar hand," that is, extension of the hand combined with flexion at the wrist. Shortly afterward there appeared clonic motions of the arm, and the animal whirled about on its ischial protuberances and fell to its right side. The clonic jerks then extended into the right leg. The stimulus was stopped after about two minutes, but the seizure persisted for a minute longer, and involved the left side of the body as well as the right side. Then for a time he lay on the left side with the left arm and leg extended, and the right arm and leg flexed. There was no deviation of the eyes, nor anisocoria, and adversive movements of the head were not noticed. Gradually he relaxed, and arose to a sitting posture, but he appeared dazed. The face was flushed, and for a few minutes the right hand hung limply as he moved about. He had voided at the beginning of stimulation.
5. Experiments with a monkey in which an electrode was inserted in the hypothalamus. On Nov. 17, 1933, an adult male Macacus rhesus weighing 6.5 kg. was operated on under sodium amytal anæsthesia, and a coil was placed in a circular opening in the calvarium. The indifferent electrode was laid beneath the scalp, and the active platinum electrode was brought forward over the surface of the frontal lobe, which was elevated to permit insertion of the wires into the tissue just anterior to the optic chiasm. The tip of the wire was pushed backwards about 1 cm. from this point into the hypothalamus.
Ten days later wound healing was complete, and the animal was stimulated for a period of ten minutes at 10 impulses per second. At first he was restless and moved about the cage, then he became quiet, and yawned several times. There was no urination, pupillary change, nor bowel movements. He remained quiet until the end of the period. Two minutes after stimulation ceased the animal became drowsy, and his head drooped. It was difficult to arouse him with a stick, although five minutes later he woke up for a short time and took interest in his surroundings. His head was soon down again, and for 5 minutes he appeared to be asleep. He was finally aroused by a noise, but not until 30 minutes after the stimulation did he become bright and active.
--------------------------------------------------------------------------------
ELECTRICAL EXCITATION 127
On a second period of stimulation the same results followed, but we noticed also that there was a general vasodilatation over the entire skin which faded out as the current was stopped, and started when it was recommenced.
In summary a method is presented for the study of excitable regions in the nervous system during the normal life of an animal. It employs two coils which are actually the primary and secondary elements of an induction coil. The secondary coil is entirely imbedded inside the animal and the primary coil is of large size and carries currents of high peak value. The animal is permitted to live comfortably within a cage through which passes the magnetic field from the primary coil. Stimulation occurs at one of the terminals of the secondary coil which can be placed in any portion of the central or peripheral nervous system. A description is given of the electrical circuits employed and several examples are cited to indicate the range of usefulness of the apparatus.
BIBLIOGRAPHY
1) Adrian, E. D.: The Mechanism of Nervous Action. Philadelphia, Univ. of Penn. Press, 1932, p. 3.
2) Beevor, A. and Horsley, V.: A minute analysis (experimental) of the various movements produced by stimulating in the monkey different regions of the cortical center for the upper limb, as defined by Professor Ferrier. Phil. Trans. Roy. Soc., London, 1887,178B, 153-67.
3) Cannon, B.: A melhod of stimulating autonomic nerves in the unanæsthetized cat with observations on the motor and sensory effects. Am. J. Physiol., 1933, 105, 366-72.
4) Cole, K. S.: Electric excitation in nerve. Cold Spring Harbor Symposia on Quantitative Biology, 1933, 1,1-13.
5) Cushing, H.: Faradic stimulation of postcentral gyrus in conscious patients. Brain, 1909, 32, 44-53.
6) DuBois-Reymond, E.: Untersuchungen über thierische Elektricität. Berlin, 1848.
7) Erlanger, J. and Garrey, W. E.: Faradic stimuli: A physical and physiological study. Am. J. Physiol., 1914, 35, 377-473.
8) Ferrier, D.: The Functions of the Brain. G. P. Putnam's Sons, N. Y., 1880.
9) Feser, J.: Drahtlose Reizung der Froschnerven mittels elektrischer Wellen. Deut. tierärtz. Wchschr., 1930, 38, 23-4.
10) Franklin, B., in a Jetter to Dr. Pringle, read before the Royal Society, Jan. 12, 1758. Quoted in part in (2), pp. 412-13. Reprinted in full in The American Museum: or Repository of Ancient and Modern Fugitive Pieces, etc., Prose and Poetical. 1789, 6, 53-4.
11) Fritsch, G. and Hitzig, E.: Ueber die elektrische Erregbarkeit des Grosshims. Archiv. f. Anat., Physiol. u. wissen. Med., Leipzig, 1870, p. 300.
--------------------------------------------------------------------------------
128 YALE JOURNAL OF BIOLOGY AND MEDICINE
12) Galvani, Luigi: Collezione delle Opera, Bologna, 1841, pp. 17-27. [See also:] Dalton, J. C.: The experimental Method in Medical Science. Lecture 1. Galvani and Galvanism in the Study of the Nervous System. N. Y., G. P. Putnam's Sons, 1882.
13) Galvani, Luigi: De viribus electricitatis in motu musculari, Commentarius. Bologna, 1791.
14) Hess, W. R.: Beiträge zur Physiologie des Hirnstammes. 1. Teil. Die Methodik der lokalisierten Reizung und Ausschaltung subkortikaler Hin abschnitte. Leipzig, 1932.
15) Keeton, R. W. and Becht, F. C.: The stimulation of the hypophysis in dogs. Amer. J. Physiol., 1915, 39, 109-22.
16) Keeton, R. W.: Personal communication to Dr. William Mahoney, April, 1934.
17) Lapicque, L., and Lapicque, Mme. L.: Sur la forme de la loi d'excitation electrique exprimee par la quantite. Reponse a M. Hoorweg. Compt. rend. Soc. de biol., 1905, 58, 668-70.
18) Loucks, R. B.: Preliminary report of a technique for stimulation or destruction of tissues beneath the integument and the establishing of conditioned reactions with faradization of the cerebral cortex. J. Comp. Psychol., 1933, 16, 439-44.
19) Lucas, K.: The conduction of the nervous impulse. London, 1917.
20) Magendie, F.: Experiences sur les fonctions des racines des nerfs rachidiens. J. de Physiol. Exper. et Path., 1822, t. ii, p. 276.
21) Marat, J. P.: Memoire sur l'electricite medicale. Paris, Méquignon, 1784.
22) Mussen, A. T.: Cerebellum and Red Nucleus: A preliminary report on a new method of physiologic investigation. Arch. Neurol. & Psychiat., 1934, 31, 1 10-26.
23) Oersted, H. C.: Experimenta circa effectum conflictus electrici in acum magneticam. Copenhagen, 1820.
24) Priestley, J.: The History and Present State of Electricity, with Original Experiments. London, 1767, p. 83.
25) Ibid., p. 146-7.
26) Ibid., p. 152.
27) Sherrington, C. S.: The integrative action of the nervous system. New Haven, Yale U niv. Press, 1906.
28) Smith, W. K.: Motor cortex of the bear (Ursus americanus). Arch. Neurol. & Psychiat., 1933,30,14-39.
29) Turrell, W. J.: Three Electrotherapists of the Eighteenth Century. Ann. Med. Hist., 1921,3,361-67.
30) Volta, A.: An account of some discoveries made by M. Galvani, with experiments and observations on them. London, Phil. Trans. Roy. Soc., 1793.
31) Wesley, J.: The Desideratum: or, Electricity made Plain and Useful. By a Lover of Mankind and of Common Sense. London, 1759.
Prenumerera på:
Inlägg (Atom)